-
Разноспоровые полиподиевые папоротники: конвергентность разноспоровости.
У разноспоровых папоротников гаметофиты раздельнополые и сильно редуцированные. Разноспоровость, т. е. дифференциация спор на микро- и мегаспоры (иногда их называют мужскими и женскими спорами), возникала в процессе эволюции, несколько раз. У одного из современных папоротников, а именно у монотипного северо-восточноавстралийского рода платизома (Platyzoma)уже давно была обнаружена начинающаяся разноспоровость, что полностью подтверждается новейшими исследованиями.
Мелкие споры при прорастании дают начало гаметофиту, на котором развиваются только антеридии. Из крупных спор развиваются гаметофиты, на которых в молодости образуются антеридии, а позже только архегонии. Таким образом, если мелкие споры можно назвать микроспорами, то более крупные споры не являются еще настоящими мегаспорами. Разноспоровость у платизомы еще не полная, не вполне установившаяся.
Биологическое преимущество разноспоровости перед равноспоровостью заключается в том, что гаметофит развивается внутри споры (становится эндоспорическим) и развивается за счет тех питательных веществ, которые содержатся в споре, особенно обильно - в мегаспоре. Благодаря быстрому развитию сильно редуцированного паразитического гаметофита процесс оплодотворения происходит рано, гораздо раньше, чем у равноспоровых папоротников. Обильная пища, накопленная в мегаспоре, создает также максимально благоприятные условия для развивающегося зародыша.
-
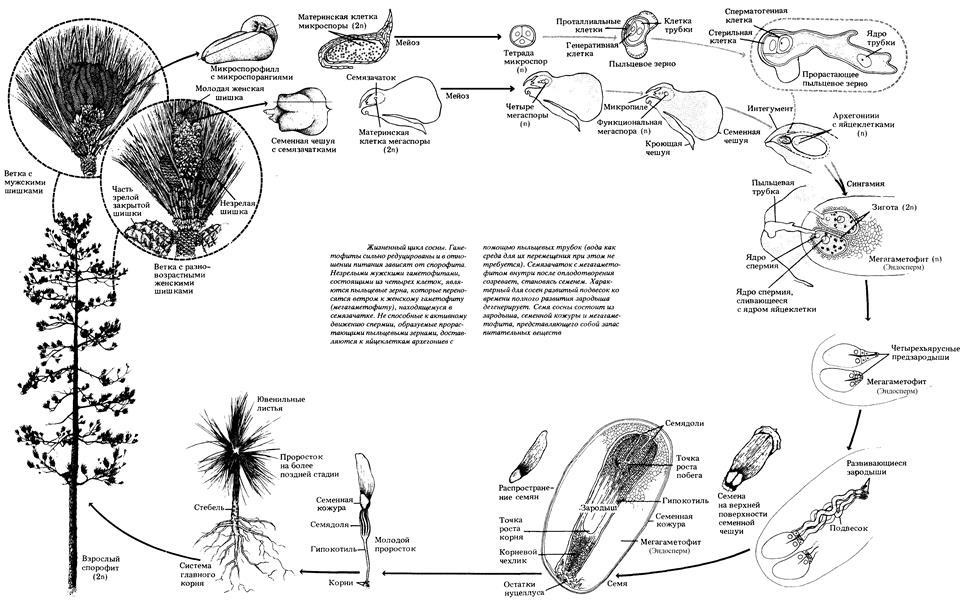
Голосеменные: общая характеристика и эволюция жизненного цикла.
Отдел голосеменных (или) в настоящее время насчитывает около 800 видов древесных и кустарниковых растений. Травянистые формы известны среди ископаемых голосеменных. Эти растения играют важнейшую роль в формировании лесных ландшафтов и накоплении биомассы и углерода, особенно в северных регионах земного шара.
Эволюция:
Голосеменные появились более 300 млн лет назад, почти на 100 млн. лет раньше первых покрытосеменных. Расцвет их приходится на значительный период, длившийся около 100–150 млн лет. Учитывая палеонтологические данные, в истории этих растений можно выделить 3 периода:
1) карбон – пермь: широко и повсеместно распространились семенные папоротники, на огромных территориях произрастали кордаитантовые растения, появились хвойные;
2) триас– юра: в этот период появились и исчезли кейтониевые и ряд ископаемых форм хвойных, широко были представлены беннетиттовые, гинкговые, цикадовые и чекановские;
3) меловой и третичный периоды: угасание гинкговых, цикадовых и чекановских, господство многих видов хвойных, особенно в Северном полушарии; к концу этого периода сложился близкий современному состав голосеменных.
Предками голосеменных, вероятно, являлись древнейшие разноспоровые папоротники, которые уже в девонский период дали начало примитивным семенным папоротникам, полностью вымершим в начале мелового периода.
Основные признаки и характеристика:
-
Имеются незащищенные семяпочки (мегаспорангие), расположенне на мегаспорофиллах, в случае редукции мегаспорофиллов семяпочки кажутся расположенными на верхушке побега.
-
Разноспоровые растения
-
В цикле развития, как и у папоротникообразных преобладает спорофит (само растение)
-
Исключительно древесные растения (деревья, кустарники, лианы)
-
Промежуточная ступень между папоротниками и покрытосеменными
-
Завязь имеет вид простой чешуи, на которой сидит одна или несколько семяпочек, иногда чешуя не развивается
В процессе развития зародыша семязачаток превращается в семя - основную единицу расселения семенных растений. У подавляющего большинства семенных растений это превращение семязачатка происходит на самом материнском растении. Но у саговниковых и у рода гинко семязачатки могут опадать еще до образования зародыша и даже до оплодотворения, и тогда оплодотворение и развитие зародыша часто происходит на земле.
В еще большей степени это относится к таким вымершим группам голосеменных, как семенные папоротники и кордатовые. Совершенно очевидно, что семена гинко и саговниковых, а тем более семенных папоротников и некоторых других вымерших групп, представляют собой более раннюю стадию эволюции семени. Но тем не менее, будь то на материнском растении или на земле, зародыш рано или поздно формируется, и в обоих случаях семязачаток превращается в семя.
Для примитивных семян, в том числе для семян саговниковых и гинкго, характерно также отсутствие периода покоя. Для большинства же семенных растений характерен более или менее длительный период покоя. Период покоя имеет большое биологическое значение, так как он дает возможность пережить неблагоприятное время года, а также способствует более далекому расселению.
В семени уже содержится, причем в очень хорошей, надежной упаковке, зародыш - крошечный спорофит с корешком, почечкой и зародышевыми листьями (семядолями).
Мужской гаметофит также претерпевает редукцию вплоть до полной потери вегетативных (проталлиальных) клеток у наиболее высокоорганизованных представителей.
В пределах отдела хорошо прослеживается появление и совершенствование пыльцевой трубки, наиболее развитой у представителей порядка хвойных.