

60. Происхождение, разнообразие, эволюция гинецея.
Рис.
1. Схема эволюции основных типов гинецея.
Апокарпный гинецей дает начало
синкарпному, от которого, в свою очередь,
происходят паракарпный и лизикарпный.
Паракарпный и лизикарпный типы изображены
в двух стадиях эволюции, что дает
представление о двух разных путях их
происхождения от исходного синкарпного
типа. Во многих случаях паракарпный
гинецей происходит непосредственно от
апокарпного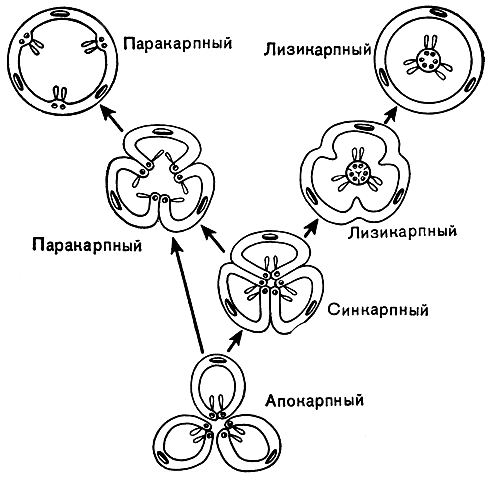
Совокупность плодолистиков в цветке называется гинецеем. Наиболее примитивные типы гинецея характеризуются свободными плодолистиками, расположенными в спиральном порядке. Гинецей, состоящий из свободных плодолистиков, например гинецей магнолии, лютика или пиона, называется апокарпным. Апокарпный гинецей характерен для относительно наиболее примитивных семейств. Число плодолистиков в апокарпном гинецее может быть очень различно - от многих до одного.
В процессе эволюции гинецея плодолистики постепенно срастаются между собой и из апокарпного гинецея возникает ценокарпный. В ценокарпном гинецее отдельные столбики (стилодии) могут оставаться свободными или же срастаются, образуя один общий столбик (сложный столбик). Ценокарпный гинецей бывает трех типов: синкарпный, паракарпный и лизикарпный.
Синкарпным называется гинецей из различного числа замкнутых плодолистиков, сросшихся между собой боковыми частями. Он дву- многогнездный и характеризуется тем, что семязачатки расположены вдоль швов замкнутых плодолистиков, т. е. по углам, образованным брюшными их частями (так называемая угловая плацентация). Хорошим примером синкарпного гинецея могут служить лилия и тюльпан.
Гораздо чаще встречается паракарпный гинецей. Для паракарпного гинецея характерна разомкнутость отдельных плодолистиков при сохранении связи краевых участков соседних плодолистиков. В отличие от синкарпного гинецея паракарпный гинецей морфологически одногнездный и характеризуется так называемой постенной или париетальной плацентацией. Общеизвестные примеры паракарпного гинецея - мак, огурец, тыква.
В некоторых линиях развития двудольных, например у портулаковых и первоцветных, из синкарпного гинецея возник лизикарпный. Это одногнездный гинецей, у которого плацентация не париетальная, как у гинецея паракарпного типа, а так называемая свободная центральная, или колончатая. Одногнездность лизикарпного гинецея возникает в процессе эволюции (или в онтогенезе) в результате исчезновения перегородок (боковых стенок) синкарпной завязи. При этом сросшиеся краевые части не разрушаются и не расходятся, и на них по-прежнему продолжают сидеть семязачатки. Таким образом, эти сохраняющиеся краевые части плодолистиков вместе с плацентами превращаются в центральную колонку, возвышающуюся в полости завязи.
61. Андроцей у покрытосеменных: происхождение, многообразие, эволюционное развитие.
Как по функции, так и по морфологической природе тычинки представляют собой разновидность микроспорофиллов. В подавляющем большинстве случаев тычинка имеет более или менее нитевидную форму и снабжена лишь одной жилкой; однако самые примитивные формы тычинок, особенно представляют собой относительно широкую лентовидную пластинку. Они еще не дифференцированы на тычиночную нить, пыльник и связник, и, таким образом, различие между фертильной и стерильной частями отсутствует или слабо выражено. Такие архаические лентовидные тычинки снабжены четырьмя попарно сближенными удлиненными линейными микроспорангиями, более или менее погруженными в ткань пластинки. Они расположены на поверхности пластинки между средней жилкой и боковыми жилками обычно трехнервных тычинок.
Совокупность тычинок в цветке называется андроцеем. У некоторых относительно более примитивных семейств тычинки сидят еще в спиральном порядке и число их неопределенное, иногда довольно большое, но у большинства цветковых растений они расположены циклически или мутовчато и число их определенное, чаще всего небольшое. Однако в некоторых линиях эволюции число тычинок может вновь возрасти, что мы видим, например, у аизооновых, кактусовых или у розы (иногда до 300 тычинок в цветке). Это увеличение числа тычинок связано с биологией опыления и чаще всего встречается у цветков, лишенных нектара. Однако в эволюции цветковых растений гораздо чаще происходит не увеличение, а уменьшение числа тычинок, и в некоторых случаях андроцей состоит лишь из одной функционирующей тычинки.
В процессе эволюции андроцея нити тычинок часто срастаются как между собой, так и с другими членами цветка. Иногда тычинки срастаются настолько тесно, что бывает даже трудно отграничить их друг от друга, что особенно хорошо выражено у тропического американского рода циклантера (Cyclanthera) из семейства тыквенных. Нити тычинок часто срастаются в пучки, как у зверобойных, или в окружающие завязь трубки, как у мальвовых. Вообще как отдельные тычинки, так и андроцей в целом характеризуются, подобно другим частям цветка, высокой эволюционной пластичностью.