

Тести з теми “Вступ до обміну речовин ”
1. При вивченні дії антибіотику олігоміцину було встановлено , що він впливає на енергетичний метаболізм . Як саме?
A.Гальмує функцію АТФ-синтетази
B.Інгібує цитохромоксидазу
C.Активує ФАД-залежні дегідрогенази
D.Активує НАДзалежні дегідрогенази
E.Посилює синтез АТФ
2. При окисленні сукцинату транспорт електронів іде за коротким дихальним ланцюгом , тобто комплекс І відсутній . Укажіть вітамін , який входить до складу цього комплексу .
A.Вітамін С
B.Нікотинамід
C.Тіамін
D.Піридоксаль
E.Фолієва кислота
3. Під час прийому снотворних препаратів - похідних барбітурової кислоти можливі прояви гіпоенергетичного стану. Активність яких ферментів тканинного дихання знижена при цьому ?
A. Цитохромоксидаза B. НАДН -дегідрогеназа
C.Малатдегідрогеназа
D.Піруватдегідрогеназа
E.Ізоцитратдегідрогеназа
4. При транспорті протонів і електронів у процесі тканинного дихання звільняється енергія . У якому процесі вона використовується ?
A.Перекисному окисленні
B.Субстратному фосфорилюванні
C.Окисному фосфорилюванні
D.Мікросомальному окисленні
5.Важливими компонентами електронного транспорту в мітохондріях є цитохроми . Який компонент цих білків бере участь в окисно -відновних реакціях ?
A.Гістидин глобіну
B.Гемінове залізо
C.Негемінове залізо
D.Білковий компонент
E.Азот пирольного циклу
51
6. При вивченні дії ціанідів було встановлено , що вони незворотньо інгібують термінальний фермент дихального ланцюга . Який саме?
A.АТФ-синтетазу
B.Цитохромоксидазу
C.ФАД-залежні дегідрогенази
D.НАДзалежні дегідрогенази
E.НАДН -дегідрогенази
7. При окисленні пірувату , малату , ізоцитрату транспорт електронів іде за довгим дихальним ланцюгом . Чому дорівнює коефіцієнт фосфорилювання для цих субстратів ?
A.1
B.2
C.3
D.4
E.5
8. У ЦТК є реакція , в якій безпосередньо синтезується 1 молекула АТФ. Назвіть її.
A.Перекисне окислення
B.Субстратне фосфорилювання
C.Окисне фосфорилювання
D.Мікросомальне окислення
9. Важливими компонентами електронного транспорту в мітохондріях є цитохроми . За якими властивостями вони розташовані у дихальному ланцюзі ?
A.За молекулярною масою
B.За зарядом молекули
C.За міцністю зв’язку з ліпідами мембран
D.За електрофоретичною рухомістю
E.За ростом окисно -відновних потенціалів
Еталони відповідей :
1- А; 2 – В; 3 – В; 4 – С; 5 – В.
52
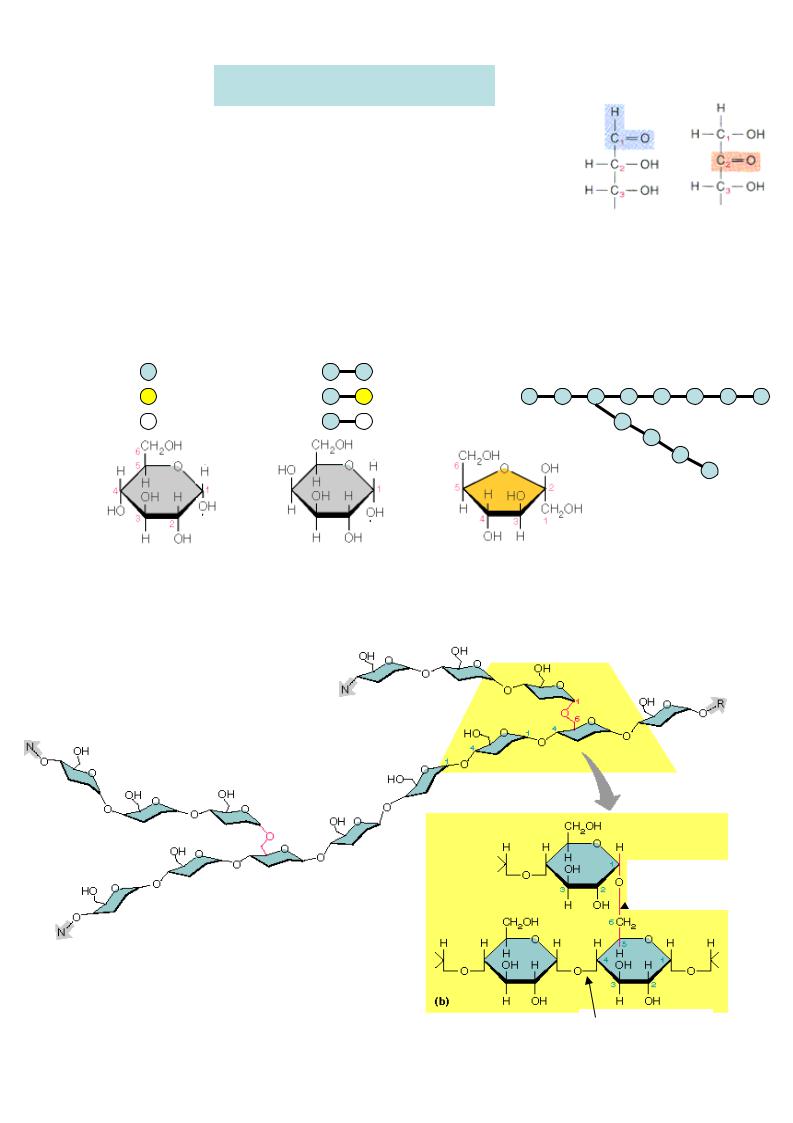
ОБМІН ВУГЛЕВОДІВ
ВУГЛЕВОДИ – це альдегідота кетопохідні
|
багатоатомних спиртів (альдози та кетози) |
|
|
||
Функції вуглеводів: |
|
|
|
|
|
енергетична (моносахариди, гомополісахариди); |
R |
R |
|||
структурна (гетерополісахариди); |
|
альдоза |
кетоза |
||
вуглеводи |
входятьдо складу складних білків – глікопротеїнів |
|
|
||
(ферментів, гормонів, рецепторів, імуноглобулінів); |
|
|
|||
вуглеводи |
використовуються у синтезі нуклеотидів, нуклеїнових кислот , |
|
|||
коферментів . |
Дисахариди: |
Полісахариди: |
|
||
Моносахариди: |
|
||||
глюкоза |
|
мальтоза |
Крохмаль (амілоза та амілопектин ) |
||
фруктоза |
|
сахароза |
глікоген |
|
|
галактоза |
|
лактоза |
целюлоза |
|
|
глюкоза |
галактоза |
фруктоза |
Крохмаль – це розгалужений гомополісахарид , с кладається з мономеру глюкоз и. Мономер и лінійних ділянок (амілози)
зв’язані 1,4-глікозидними зв’язка-
ми, а в точках розгалуження – 1,6-
глікозидними зв’язками.
Глікоген – це полісахарид , його структура подібна крохмалю, але глікоген більш розгалужений . Він містить більше 1,6-глікозидних зв’язків і більшу кількість кінцевих мономер ів, що полегшує його розпад.
1,6-глікозидний  зв’язок
зв’язок
1,4-глікозидний |
53 |
зв’язок |

Головні процеси в обміні вуглеводів
1.Травлення вуглеводів
2.Синтез глікогену - глікогенез
3.Розпад глікогену - глікогеноліз
4. Дихотомічний розпад глюкози – непряме окиснення глюкози при розпаді її молекули на дві тріози:
4.1. Анаеробний гліколіз – окиснення глюкози при відсутності кисню з утворенням 2 молекул лактату та 2 молекул АТФ. 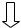
4.2. Аеробний гліколіз – окиснення глюкози при наявності кисню з утворенням 2 молекул пірувату та 8 молекул АТФ.
4.3. Спиртов е бродіння – п еретворення глюкоз и в етіловий спирт п ід дією мікроорганізмів (дріжджів ).
5.Глюконеогенез - синтез глюкоз и з не вуглеводів: ам інокислот (білків), гліцерину (ліпідів), пірувату, лактату.
6.Апотомічний розпад глюкози - пряме окиснення глюкози – пентозофосфатний шлях окиснення глюкози 
7.Взаємоперетворення моносахарів – включення інших гексоз в обмін глюкози
Вуглеводи їжі |
|
|
глюкоза |
фруктоза |
галактоза |
глікоген 
 глюкозо -6-фосфат
глюкозо -6-фосфат
ліпіди |
|
|
|
СО2 |
гліцерин |
3-фосфогліцеральдегід |
рибозо - |
||
|
|
|
|
|
|
|
|
|
5-фосфат |
білки |
амінокислоти |
піруват |
лактат |
НАДФН+ |
|
СО2 |
ацетил -SКоА |
етанол |
|
|
|
|
||
|
СО2 |
ЦТК |
|
нуклеотиди |
|
АТ |
дихальний ланцюг |
ліпіди |
коферменти |
|
Ф |
ДНК, РНК |
||
54 |
|
Н2О |
|
|
1. Травлення вуглеводів
Травлення вуглеводів – це гідроліз олігота полісахаридів, тобто розщеплення в присутності води глікозидних зв’язків під дією гідролітичних ферментів.
Ротова порожнина
Травлення вуглеводів починається у ротовій порожнині, в котрій діє α-амілаза слини (рН opt = 6,7). Фермент каталізує ідролізг 1,4-глікозидних зв’язків у молекул і крохмалю. Цей фермент є ендоамілазою, тому що він розщеплює зв’язки, віддалені від кінця ланцюгу. В результат і утворюються олігосахариди (декстрини) та невелика кількість дисахариду мальтози.
α-амілаза
слини
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1,6- |
|||
|
|
|
|
|
|
|
глікозидаза |
|
У шлунку (рН 1,5-2) |
|
|
|
|||
|
травлення вуглеводів не |
|
|
||||
|
відбувається |
|
|
|
|
|
|
|
Тонкий кишечник |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
панкреатична |
|
|
|||
|
|
|
|
|
|||
|
|
|
α-амілаза |
|
|
||
|
|
|
|
|
|
|
|
У верхньому відділі тонкого кишечникадіє панкреатична α-амілаза, котра гідролізує
1,4-глікозидні зв ’язкі шляхом послідовного відщеплення |
молекул мальтоз и |
від |
декстринів. 1,6-глікозидні зв’язкі в декстринах гідролізують |
1,6-глікозидази. |
|
На дисахарид и в кишечник у діють відповідні ферменти |
(мальтаза , сахараза |
та |
лактаза ), що розщеплюють дисахариди до моносахаридів: |
|
|
мальтоза
мальтаза
сахароза |
сахараза |
|
глюкоза + глюкоза |
+ |
глюкоза + фруктоза |
+ |
лактоза |
лактаза |
глюкоза + галактоза |
+ |
55 |
|
Перетворення глюкози у тканинах |
|
Перша реакц |
ія п |
еретворення |
||||||||
|
|
|
|
|
|
|
|
глюкози у тканинах – ак-тивація |
|||||
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
глюкози |
|
|
шляхом |
||
|
|
АТФ |
АДФ |
|
фосфорилювання за участю АТФ. |
||||||||
|
|
|
У багатьох тканинах її к |
аталізує |
|||||||||
|
|
|
|
|
|
|
|
гексокіназа, а в кл ітинах печінки |
|||||
|
|
|
|
|
|
|
|
– і глюкок |
іназа. |
Глюкокіназа |
|||
|
|
|
гексокіназа |
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
специфічна для глюкоз |
и та |
||||||
|
|
|
глюкокіназа |
|
|||||||||
|
|
|
|
працює |
при в |
|
исоких |
||||||
глюкоза |
|
глюкоза -6-фосфат |
|
||||||||||
|
|
|
|
|
|
|
|
концентраціях субстрату (КM = = |
|||||
Ця реакція «зачиняє» глюкозу у клітинах, тому |
|
12ммоль /л), а |
гексокіназа |
||||||||||
що клітинні мембрани |
непрони кні для |
|
фосфорилює різні |
гексоз и та |
|||||||||
фосфорильованої глюкози. |
|
|
|
|
інгібується глюкозо -6-фосфа-том |
||||||||
|
|
|
|
|
|
|
|
(КM = 0,1ммоль /л). |
|
|
|||
2. Синтез глікогену - глікогенез
Глікоген – це тваринний гомопол ісахарид , резервна форма глюкози. Головним чином
він депонується у печінці та скелетн их м ’язах. Глікоген тварин, |
як і |
амілопектин |
|||||||||||
рослин, є розгалуженим |
полімером глюкози, в котрому залишки |
глюкози з’єднані α |
|||||||||||
(1→4)-глікозидними зв’язками. |
У точках |
розгалуження зв ’язки |
|
знаходяться у |
|||||||||
положенні α(1→6) при |
близно у |
кожного |
|
10-го |
залишка глюкози . |
Розгалужена |
|||||||
структура |
створює велику кількість кінцевих мономер ів, |
що сприяє |
оботір |
||||||||||
ферментів, |
відщеплюючих або |
приєднуючих глюкозу при |
синтез |
і або |
розпаді |
||||||||
глікогену. |
|
|
|
|
|
|
УТФ |
ФФ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
фосфо - |
|
УДФ-глюкозо - |
|
|
|
|
|||||
|
глюкомутаза |
|
|
пірофосфорилаза |
|
|
|
|
|||||
глюкоза -6-фосфат |
|
глюкозо -1-фосфат |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
УДФ-глюкоза |
|
||
|
Зберігання |
в орган |
ізмі вільної |
|
|
глюкози неможливо із-за її високої |
|||
|
розчинності: в еликі концентрац ії |
|||
|
глюкози створюють |
у |
клітинах |
|
|
гіпертонічне середови -ще, що |
|||
|
сприяє |
надходженню води. |
||
|
Навпаки , нерозчинний |
глі-коген |
||
56 Гранули глікогену |
є осмотично малоактив -ним і |
|||
компактним. |
|
|
|
|
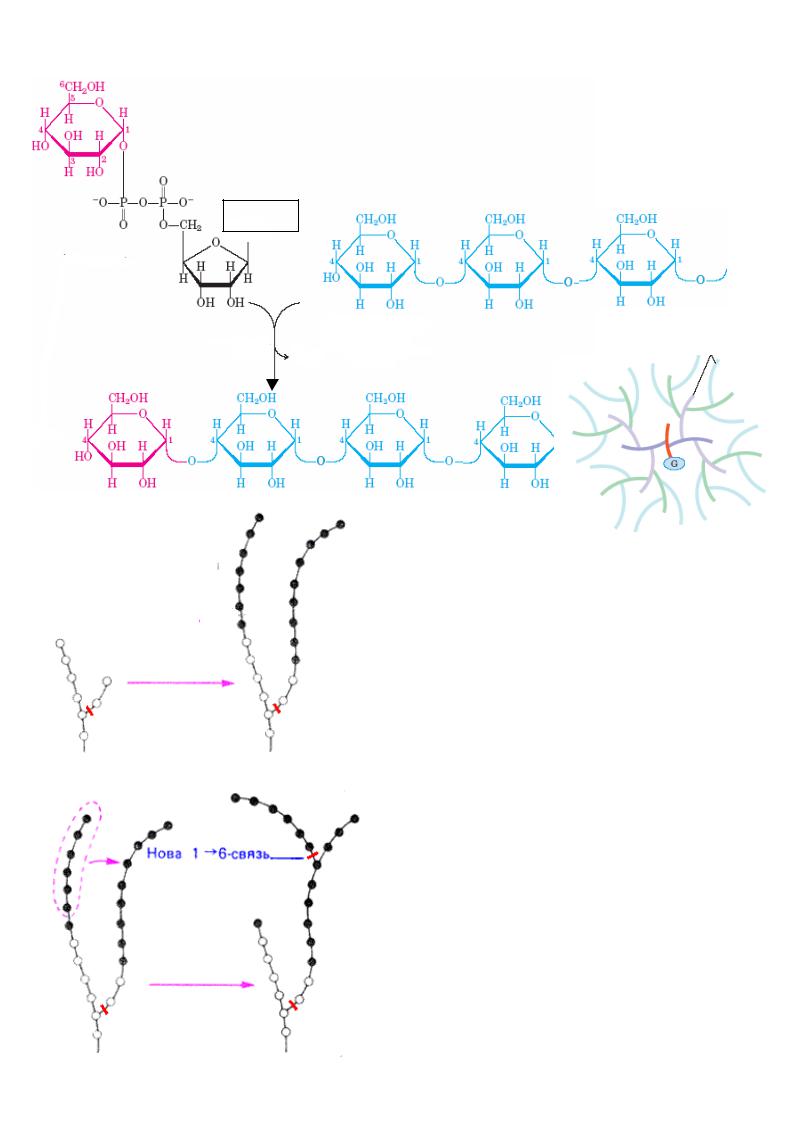
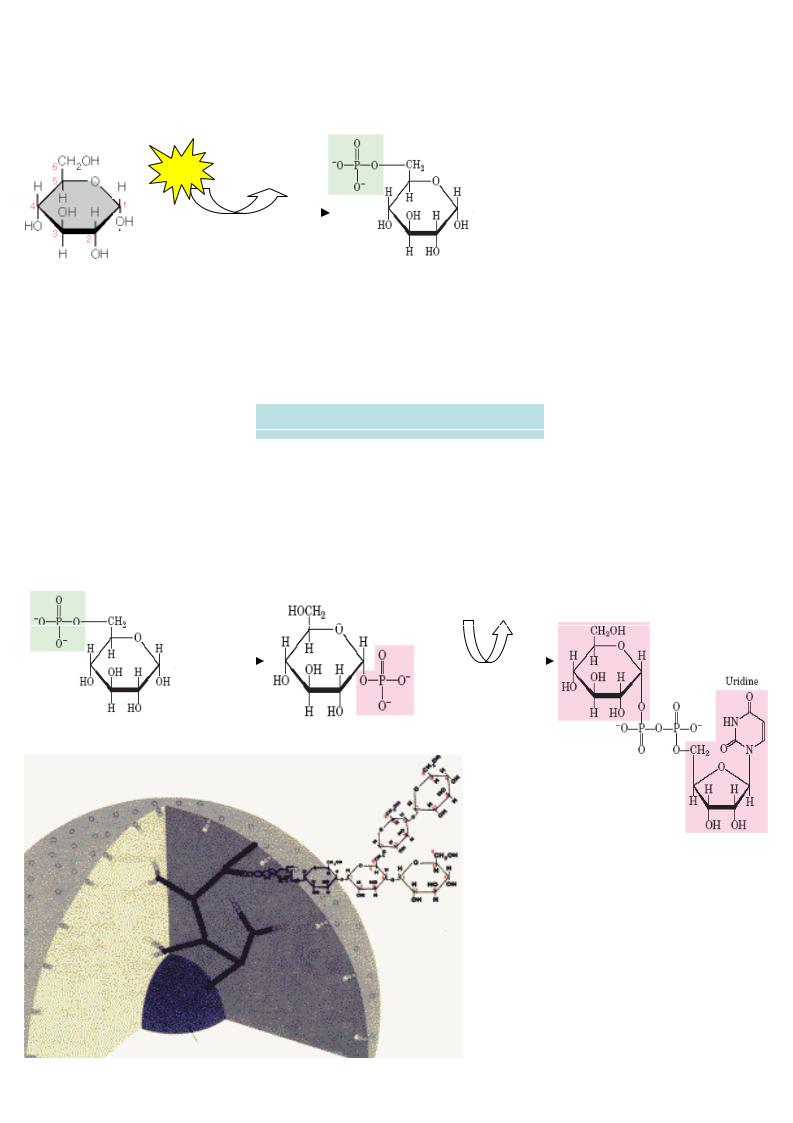
Подовження ланцюгулінійних полімерів здійснюється шляхом утворення 1,4- глікозидних зв’язків
урацил
УДФ-глюкоза
|
глікоген- |
|
олігосахарид -«запал» глікогену (праймер ), що містить n |
|
|
||
|
|
молекул глюкози (n>4) |
|
|
синтаза |
|
УДФ |
|
|
|
|
|
|
|
|
глікоген, що містить (n+1) молекул глюкози
а) подовження ланцюгу
глікогенсинтаза
б) розгалуження
фермент розгалужень
Подовження ланцюгу |
каталізується |
|||
глікогенсинтазою. |
У зв ’язку з тим, що |
|
||
утворення глікозидних зв’язків |
між сахара- |
|||
ми є ендергонічною реак цією, |
спочатку |
|||
утворюється активований попередник |
- |
|||
УДФ-глюкоза |
внаслідок |
|
реакц |
ії |
глюкозо -1-фосфату |
з уридинтрифосфатом |
|||
(УТФ). Лише після цього залишок |
глюкози |
|||
легко переносит ься з УДФ-глюкоз и на вже існуючий праймер глікогену. Ко ли ланцюг досягає довжини більш 11 залишків глюкози, спеціальний фермент розгалужень
глікогену (1,4→1,6-глікозил -трансфераза)
каталізує перен |
есення |
кінцевого |
олігосахариду, |
що містить |
6-7 залишків |
глюкози, на 6(ОН) залишок глюкоз и того ж самого або іншого ланцюгу глікогену з утворенням точки розгалуження [α(1→6)-
зв’язки]. |
Далі |
за допомогою |
глікогенсинтази цей фрагмент |
||
подовжується . |
57 |
|
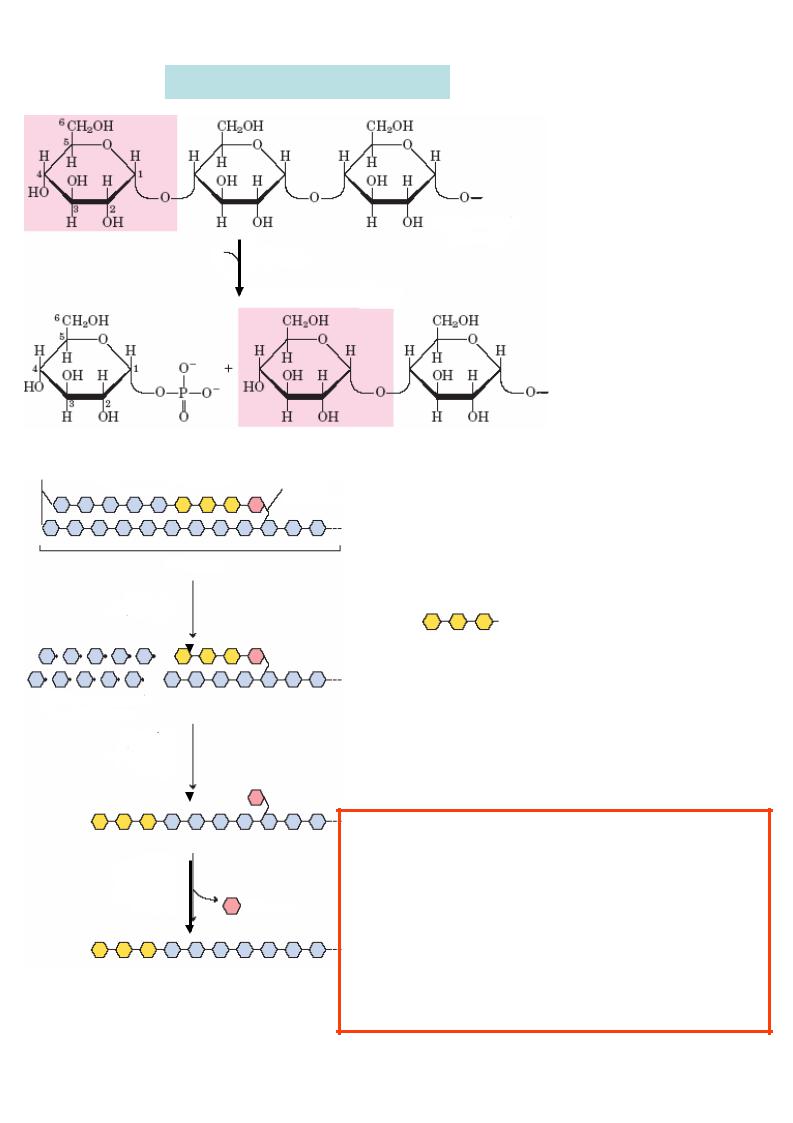
3. Розпад глікогену у тканинах
Н3РО4 Глікоген ( n молекул глюкози)
глікогенфосфорилаза
Глікоген печінки ніколи повністю не розпадаєть - ся. Скорочуються лише невідновлювальні кінці деревоподібної структу - ри глікогену шляхом фосфоролізу , тобто взаємодії з неорганічним фосфатом у місці розривання зв’язку.
|
|
|
|
|
Глікогенфосфорилаза |
|||||
|
|
|
|
|
відщеплює від |
нероз- |
||||
|
|
|
|
|
галужених ланцюгів |
|||||
|
|
|
|
|
глікогену по одному |
|||||
Глюкозо -1-фосфат |
Глікоген ( (n –1) молекул глюкози) |
мономеру |
у |
вигляді |
||||||
глюкозо -1-фосфату. |
||||||||||
|
|
|
1,6-зв’язок Глікогенфосфорилаза |
|
||||||
|
|
|
|
каталізує фосфорол із 1,4- |
||||||
|
|
|
зв’язків, тобто розщеплення |
лінійних полімерів. |
||||||
|
|
|
Розрив 1,6-зв’язків |
каталізує фермент дерозга - |
||||||
глікоген |
лужень. Фермент має дві активності : |
|
|
|||||||
1) глікозилтрансферазну активн ість: він |
||||||||||
|
|
|
||||||||
глікоген- |
|
|
переносить олігосахарид |
із тр |
ьох |
залишків |
||||
фосфорилаза |
|
|
глюкоз |
|
на кінець нерозгалуженого |
|||||
|
|
|
ланцюгу , але залишається |
одна молекула |
||||||
|
|
|
глюкози, зв’язана 1,6- глікозидним зв’язком; |
|||||||
молекули глюкозо -1-фосфату |
2) 1,6-глікозидазну активн істю: гідролізує 1,6- |
|||||||||
зв’язки з утворенням глюкоз и |
та |
лінійного |
||||||||
фермент |
|
|
||||||||
|
|
полімеру, на |
|
котрий дал |
|
і |
діє |
|||
|
|
|
||||||||
дерозгалужень |
|
|
|
|
||||||
|
|
глікогенфосфорилаза. |
|
|
|
|
|
|
||
глікозил- |
|
|
|
|
|
|
|
|
||
трансферазна |
|
|
|
|
|
|
|
|
|
|
активність |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
фермент
дерозгалужень
1,6-глікозидазна активність глюкоза
Лінійний полімер - субстрат для подальшої дії ікогенглфосфорилази.
Розпад гл ікогену в печінці та м ’язах
відрізняється лише за однією реакцією .В печінці є фермент глюкозо -6-фосфатаза, що каталізує відщеплення фосфату від глюкоз о-6-фосфату - своєрідного якоря, котрий не випускає глюкозу із клітин. Мобілізація глікогену печінки
підтримує сталу концентрац ію глюкоз и в крові. Функція гл ікогену м’язів – забезпечення м’язів метаболічним паливом.
58
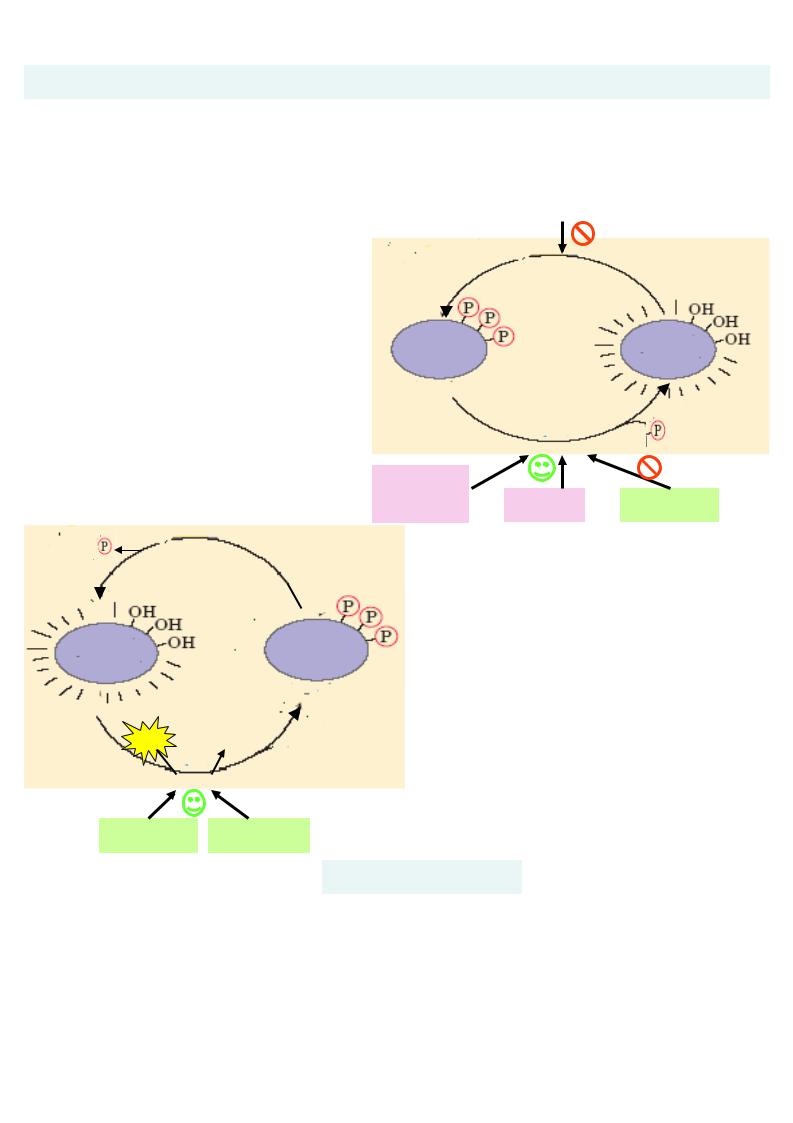
Регуляція глікогенезу та глікогенолізу (синтезу та розраду глікогену)
Синтез і розпад глікогену – це два протилежних процеса , котрі ніколи не відбуваються в клітинах одночасно . Регуляція цих метаболічних шляхів здійснюється завдяки змінам
активності |
двох регуляторних ферментів синтезу та розпаду глікогену - |
||
глікогенсинтази та глікогенфосфорілази за допомогою |
|
|
|
інсулін |
|
||
Їхнього фосфорилювання та дефосфорилювання . |
|
||
Синтез глікогену активується в період травлення (1-2 години після при йому
вуглеводної їжі). |
За цей |
час |
збільшується секреція |
гормон |
у |
підшлункової залози інсуліну, котрий активує глікогенсинтазу шляхом її дефосфорилювання . Внаслідок цього глюкоза після всмоктування у кров із кишечника депон ується у печінці у вигляді глікогену .
протеїнфосфатаза
Н2О
глікоген- |
глікоген- |
фосфорилаза |
фосфорилаза |
неактивна |
активна |
|
АТФ АДФ
протеїнкиназа
глюкагон адреналін
глікоген- |
глікоген- |
синтаза |
синтаза |
неактивна
|
Н2О |
активна |
|
|
|
|
протеїнфосфатаза |
|
глюкозо - |
інсулін |
глюкагон |
6-фосфат |
||
Розпад |
|
( мобілізація) гл |
ікогену |
||
відбувається, г оловним чином , |
у період |
||||
між прийомами їжі , а так ож під час |
|||||
фізичної |
роботи. У постабсорбтивному |
||||
періоді |
(при |
зниженні концентрац |
ії |
||
глюкози в кров |
і) підсилюється секреція |
||||
іншого гормону підшлункової залози |
- |
||||
глюкагону, |
кот рий активує |
глікоген- |
|||
фосфорилазу |
шляхом фосфорил ювання. |
||||
За цих умов у печінці відбувається розпад
гліконену , і глюкоза |
потрапляє у кров . |
Аналогічно у м ’язах |
діє адреналін. У |
скелетних м ’язах |
розпад глікогену |
активується при фізичній роботі.
Глікогенові хвороби
Це спадкові захворювання – ензимопатії, пов’язані з порушеннями активності ферментів синтезу або розпаду глікогену .
Глікогенози – хвороби накопичування глікогену, в наслідок яких спостерігаються
гіпоглікемія та її наслідки . Наприклад , хвороба Гірке характеризується |
недостатністю |
глюкозо -6-фосфатази. |
|
Аглікогенози – порушення синтез у глікогену, котрі призводять до |
зниження його |
вмісту у тканинах , а також гіпогликемії. |
59 |

|
|
|
|
глюкоза |
|
|
|
|
|
|
|
4. Дихотомічний розпад глюкози - |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
непряме окиснення глюкози. |
|||
|
|
|
|
|
|
|
|
|
гліколіз |
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
Незалежно від умов, дихотомічний |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
розподіл 6-ти |
вуглецевої молекули |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
2 піруват |
|
|
|
|
|
|
||||||||||
анаеробні умови |
|
|
|
анаеробні |
|
глюкози |
з наступним утворенням |
|||||||||||||
у дріжджях |
|
|
|
|
|
|
аеробні |
|
|
умови |
|
двох |
тривуглецевих молекул |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
пірувату |
відбувається однаково . |
||||||
2 етанол +2СО2 |
|
|
|
|
|
|
|
|
умови |
|
2 лактат |
|||||||||
|
|
|
|
|
|
|
2СО2 |
|
Далі, в залежності від умов, |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
піруват за різними шляхами може |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
2 ацетил -КоА |
|
|
|
|
|
перетворюватися в лактат |
або в |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ацетил -КоА,або в етиловий спирт |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
4СО2 + Н2О |
|
|
|
|
|
|
(в дріжджях). |
|
|
|||||||
|
|
|
|
|
|
|
|
|
4.1. Анаеробний гліколіз |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
Анаеробний гл іколіз |
– це ферментативн ий |
розпад глюкоз и до молочно ї |
кислоти |
|||||||||||||||||
(лактату) без споживання кисню (схема |
на наступній сторінці). Усі реакц ії гліколізу |
|||||||||||||||||||
локалізовані у цитозолі. |
|
м’язах у перші хвилини інтесивної роботи , |
||||||||||||||||||
Анаеробний гліколіз відбувається у |
|
|||||||||||||||||||
еритроцитах (в них |
немає мітохондрій), |
у різних органах при нестачі кисню |
||||||||||||||||||
(гіпоксія), а так ож у |
клітинах пухлин. |
За цих умов гліколіз є |
єдиним способом |
|||||||||||||||||
отримання енергії |
|
для синтезу АТФ із АДФ і Фн. |
|
|
|
|||||||||||||||
1 стадія - енергоспоживальна, в котрій глюкоза двічі фосфорилюється (активується) з витратой 2 молекул АТФ і дихотомічно розподіляється на два фосфорил ьованих С3 фрагменти - фосфотр іози: фосфод іоксіацетон и 3-фосфогл іцериновий альдег ід, які здатні взаємоперетворюватися . Але у другу стадію безпосередньо включається лише 3-фосфогліцериновий альдегід.
2 стадія - енергогенеруюча стадія (гліколітична оксидоредукція), в якій відбувається окиснення (дегідрування ) 3-фосфогліцеральдегіду та відновлення НАД+ у НАДН+Н+. Крім того, у другій стадії відбуваються дві реакції субстратного фосфорилювання. В
кожній з них утворюється по 2 молекули АТФ |
у перерахунку н а одну молекулу |
|
глюкози. Внаслідок |
дихотомії утворилися дві |
фосфотріози, кожна з них далі |
перетворюється в піруват . Таким чином, енергетичний вихід другої стадії складає 4
молекули АТФ. |
|
|
НАДН+Н+, |
що утворився внаслідок |
|||||||
|
|
|
|
|
|
|
|
|
|
||
|
|
глюкоза |
|
|
|
|
|
окиснення |
3-фосфогліцеринового альдегіду, |
||
|
|
|
|
|
|
|
|
|
|
відновлює піруват до лактату . тому що за |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
анаеробних умов він не може окиснюватися у |
|
|
|
|
|
2НАД+ |
|
|
|
||||
|
|
|
|
|
|
|
мітохондріальному дихальному ланцюзі . |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2НАДНН+ |
|
|
|
|
|
|
||
2 піруват |
|
2 лактат |
|
|
|
||||||
|
у відсутності О2 |
|
|
|
|||||||
|
Під час перетворення глюкози у лактат |
||||||||||
|
|
|
|
|
|||||||
60 |
|
|
|
|
|
|
|
|
|
утворюється 2 молекули АТФ ((2х2)-2=2). |
|
|
|
|
|
|
|
|
|
|
|
|
|