

1971 г. Алексей Матвеевич Оловников
ГИПОТЕЗА МАРГИНОТОМИИ - механизм работы счетчика кл. делений
при матричном синтезе полинуклеотидов ДНК-полимераза
не в состоянии полностью воспроизвести линейную матрицу, реплика получается всегда короче в ее начальной части.
при каждом делении клетки ее ДНК укорачивается, что, ограничивает пролиферативный потенциал клеток
сотрудник Института биохимической физики РАН,
Фото с сайта moikompas.ru
1971 г. А.М.Оловников
ВО ВСЕХ СОМАТИЧЕСКИХ КЛЕТКАХ ОРГАНИЗМА МЕХАНИЗМ РЕПЛИКАЦИИ ТЕЛОМЕР ОТСУТСТВУЕТ;
ПОЭТОМУ ПРИ ДЕЛЕНИЯХ КЛЕТОК ТЕЛОМЕРЫ ПОСТЕПЕННО УКОРАЧИВАЮТСЯ
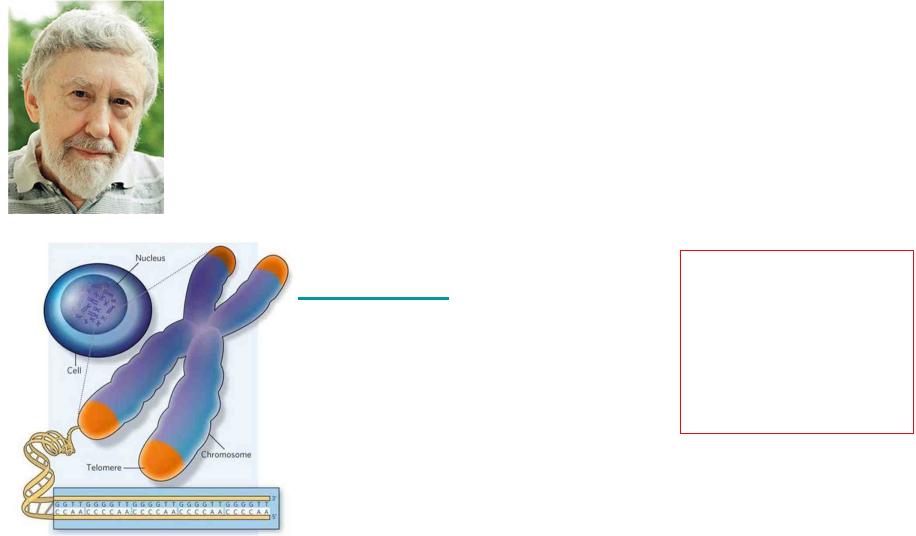
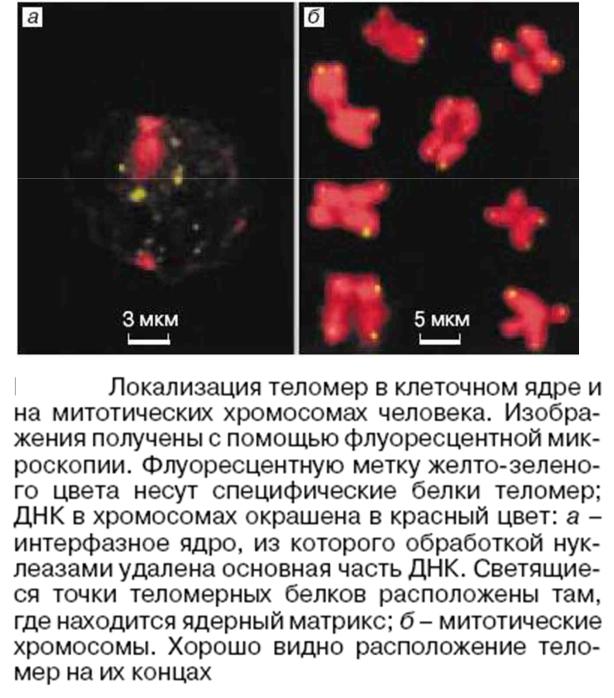
Теломеры — концевые участки хромосом, состоящие из повторяющихся последовательностей нуклеотидов
Термин теломера
(от греч. telos —
конец и meros — часть) предложил Герман Джозеф Меллер в 1932 г.
предположил существование фермента
в 1985 Грейдер и Блэкберн
ТЕЛОМЕРАЗА - фермент,
достраивающий концы линейных молекул ДНК хромосом (теломеры)
короткими повторяющимися последовательностями в половых клетках и клетках опухолей, обеспечивая их бессмертие
НОБЕЛЕВСКАЯ ПРЕМИЯ ПО ФИЗИОЛОГИИ ИМЕДИЦИНЕ- 2009
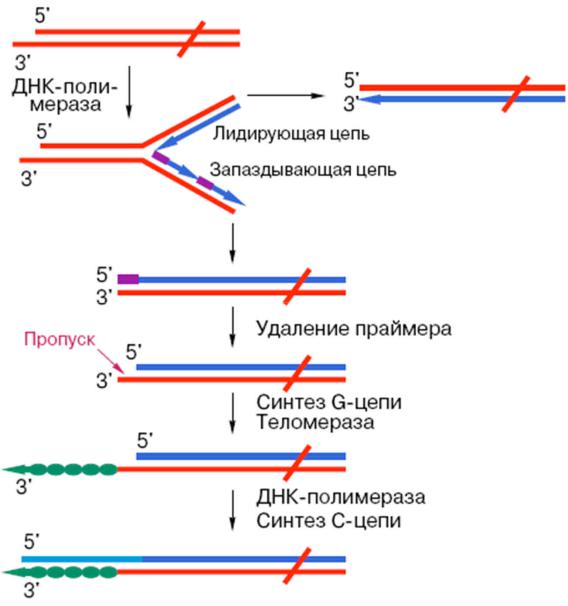
Возникновение
недореплицированного 5`конца линейной хромосомы
Синтез на этом концевом участке теломерной ДНК с помощью теломеразы
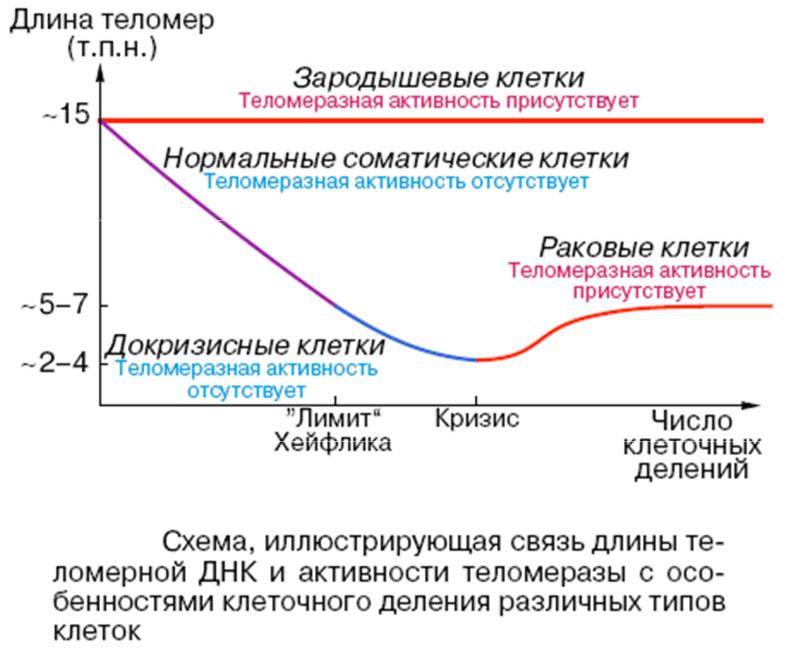
1. Концы линейных хромосом с 3'-конца ДНК заканчиваются повторяющимися последовательностями нуклеотидов, теломерами,
которые синтезируются рибонуклеиновым ферментом теломеразой
2. Половые клетки и клетки опухолей имеют высокоактивную теломеразу, которая достраивает 3'-конец ДНК, на котором реплицируется комплементарная цепь при делении.
3. Соматические клетки эукариотов, имеющие линейные хромосомы, лишены теломеразной активности. Их теломеры укорачиваются как в
процессе онтогенеза и старения in vivo), так и при культивировании in vitro.
4.Структуры теломер различаются среди простейших, однако у всех позвоночных они одинаковы (TTAGGG)n
5.Репрессия теломеразы определяет
клеточное старение в культуре (лимит Хейфлика).
6. Клетки больных синдромом преждевременного старения Хатчинсона-Гилфорда
и синдромом Дауна имеют укороченные теломеры.

|
|
Предполагае- |
|
|
|
|
мые "часы" |
Теория, гипотеза |
|
|
|
старения |
|
|
|
|
|
|
|
|
|
|
Теории "катастрофы ошибок", |
|
|
|
ДНК |
накопления спонтанных мутаций, |
|
|
|
|
маргинотомии или теломеразы |
|
|
|
|
|
|
|
|
Митохондрии |
Свободно-радикальная теория |
|
|
|
|
|
|
|
|
Клетка |
"Лимит Хейфлика" |
|
|
|
|
|
|
|
|
|
|
|
|
|
Эпифиз |
Мелатонин как счетчик внутреннего |
|
|
|
времени в солнечных часах старения |
||
|
|
|
||
|
|
|
|
|
|
|
Половые |
Инволюция половой функции |
|
|
|
железы |
||
|
|
|
|
|
|
|
|
|
|
|
|
Иммунная |
Иммуностарение |
|
|
|
система |
||
|
|
|
|
|
|
|
|
|
|
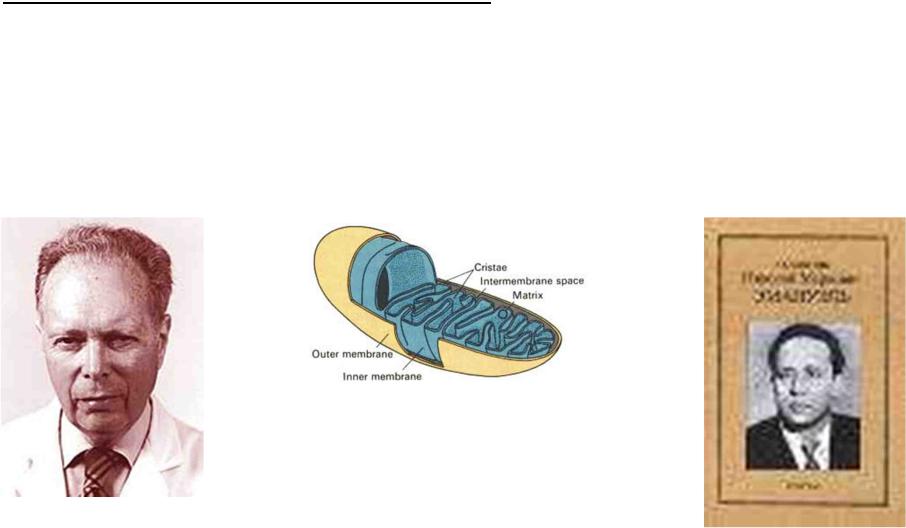
Свободнорадикальная теория старения
D.Harman в 1956 г. и Н.М.Эмануэль в 1958 г.
пpодуциpуемые главным образом в митохондриях клеток активные формы кислорода (АФК)
повpеждают клеточные макpомолекулы (ДНК, белки, липиды)