

Практические занятия / Дополнительные материалы / Биология и генетика / Мутации
.docМутационная теория (основы заложил Гуго де Фриз, 1901-1903 гг.) Современная мутационная теория содержит следующие положения:
-
Мутации - дискретные (прерывистые) изменения наследственного материала организмов.
-
Мутации возникают внезапно, в отличие от модификаций, они не образуют непрерывных рядов изменчивости, не группи-
руются вокруг среднего типа.
3. Мутации - редкие события; вероятность их возникновения составляет 1 на 10-1000 тысяч копий одного гена.
-
Одни и те же мутации в определенных условиях могут возникать неоднократно, накапливаясь в генофонде популяций.
-
Мутации не имеют направленного характера.
-
По воздействию на популяции мутации могут быть полезными, нейтральными, вредными (сублетальными) и летальными; чаще всего они снижают выживаемость мутантов.
7. Мутации могут передаваться по наследству, влияя, таким образом, на ход преобразовательных процессов.
Классификации мутаций
|
Группы мутаций |
Характеристика |
|
А. По характеру изменения генетического материала: |
|
|
1. Генные |
Изменение структуры генов |
|
2. Хромосомные |
Изменение структуры хромосом |
|
3. Геномные |
Изменение числа хромосом |
|
Б. По проявлению в гетерозиготе розиготе |
|
|
1. Доминантные |
Проявляются в гетерозиготном состоянии |
|
2. Рецессивные |
Не проявляются в гетерозиготном состоянии |
|
В. По уклонению от нормы (от дикого типа): |
|
|
1. Прямые |
Изменение обычного состояния генетического материала |
|
2. Обратные (реверсии) |
Возврат к исходному состоянию гена. |
|
Г. В зависимости от причин, вызывающих мутации: |
|
|
1. Спонтанные |
Возникают без видимой причины, т.е. без каких-либо индуцирующих воздействий со стороны экспериментатора, вследствие нарушения процессов репликации и репарации |
|
2. Индуцированные |
Возникают под действием мутагенных факторов (химических, физических и биологических мутагенов) |
|
Д. По локализации в клетке |
|
|
1. Ядерные |
Мутации в генах, локализованных в ядерном геноме |
|
2. Цитоплазматические |
Мутации в генах митохондрии и пластид |
|
Е. По отношению к возможности наследования |
|
|
1. Генеративные |
Происходят в половых клетках, передаются по наследству |
|
2. Соматические |
Происходят в соматических клетках |
|
Ж. По значению (по воздействию на популяции) |
|
|
1. Полезные |
Повышают адаптацию к условиям среды |
|
2. Нейтральные |
Не влияют на адаптацию к условиям среды |
|
3. Сублетальные |
Снижают жизнеспособность |
|
4. Летальные |
Приводят к гибели организма |
Хромосомные мутации - хромосомные перестройки (аберрации) (от лат. aberratio - уклоняться), изменяющие их структуру. Хромосомные мутации, или перестройки, характеризуются изменениями положения участков хромосом. Выделяют внутрихромосомные перестройки (делеции, дупликации, инверсии), когда изменяется одна хромосома, или хромосомы одной гомологичной пары; и межхромосомные аберрации (транслокации), когда в перестройки вовлечены участки разных негомологичных хромосом.
Делеция - потеря какого-либо участка хромосомы - промежуточного или концевого. Делеции одних и тех же локусов в обеих гомологичных хромосомах обычно легальны, так как утрачивается значительный объем генетической информации. Таким образом, делеции возникают вследствие потери хромосомой того или иного ее участка.
Дупликация (повторение) - присутствие одного и того же участка хромосомы более чем в одном экземпляре в одной хромосоме или в разных негомологичных хромосомах. При дупликации, в отличие от делеции, происходит удвоение участка хромосомы.
Многие дупликации вызывают появление новых фенотипических признаков, не снижающих жизнеспособность организма. Участки с высокой и умеренной повторностью нуклеотидных последовательностей имеются в генотипах многих млекопитающих и других классов животных и растений. Например, около 10% генома мыши составляют тандемно повторяющиеся до 10 раз нуклеотидные последовательности, содержащие по 10 нуклеотидов. Дупликации многих генов повышают устойчивость организма к различным мутациям и увеличивают генетическое богатство популяции.
Многие дупликации и делеции возникают в результате разрывов хромосомы вследствие действия ионизирующей радиации, химических веществ или вирусов. Они могут также возникать при неравном кроссинговере, когда конъюгация гомологов происходит неточно, если в соседних участках хромосом находятся сходные последовательности ДНК.
Инверсия - поворот отдельного участка хромосомы на 180°; при этом число генов в хромосоме остается прежним, а изменяется лишь их последовательность. Инверсия может возникнуть при образовании хромосомой петли с последующим разрывом ее основания и растяжением петли в стороны. Таким образом, инверсией обозначают поворот участка хромосомы на 180 градусов.
Инверсия, действуя как «ингибитор кроссинговера», может нарушить процесс конъюгации во время мейоза и привести к гибели гамет. Если этого не происходит, в фенотипе развивающегося из зиготы организма возникают изменения, как следствия зависимости действия генов от их последовательности в хромосоме (эффект положения).
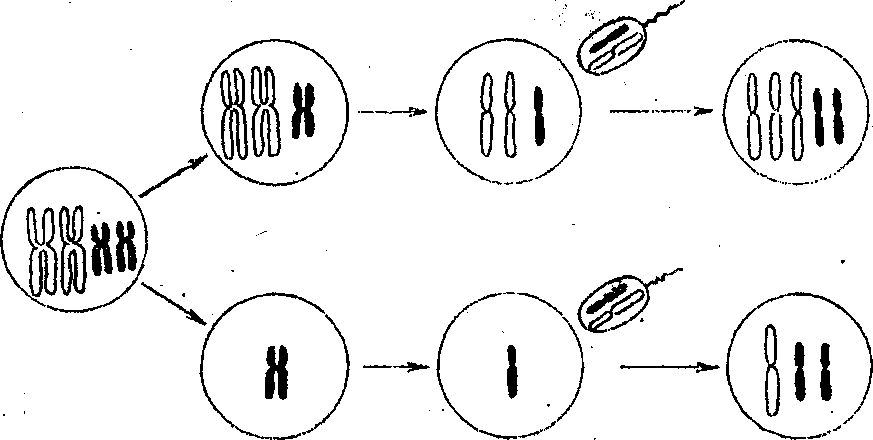
|
Фенотипически генотипически нормальный N=46 |
Фенотипичес нормальный носитель транслокации N=45
|
Синдром Дауна N=46 |
Трисомик но хромосоме N=46 (не выявлены) |
|
Моносомяк по хромосоме 14, N=45 (не выявлены) |
|
Моносомик но хромосоме 21 , N=45 (не выявлены) |
|
|
|||||||
|
|
Гаметы
хема формирования гамет (I) и зигот (II) при нерасхождении пары хромосом в первом мейоти-ческом делении: А - трисомия, Б -моносомия в зиготе.
Фенотипически
нормальный
носитель
транслокации
(N
=
45 )
Мейоз
Нормальные
гаметы
партнера
Рис.
21.28. Синдром Дауна обусловленный
транслокацией
хромосом 14-21. В потомстве фенотипически
нормального носителя транслокации
(имеющего 45 хромосом в наборе)
могут
быть дети Нормальные фенотипически
генотипически
нормальные фенотипически,
но
явияющиеся ноcителями
перестройки,
и, наконец, больные с синдромом Дауна.
![]()








При умножении геномов диплоидных гибридов образуются аллополиплоиды (от греч. allos - другой, иной). При этом происходит восстановление плодовитости гибрида.
Часто отдаленные гибриды оказываются бесплодными. Так, например, редечно-капустный гибрид имеет в своих клетках 18 хромосом, которые образованы двумя разными геномами: R (редьки) и В (капусты), каждый из которых представлен девятью хромосомами. В мейозе эти негомологичные хромосомы не могут конъюгировать между собой. Они беспорядочно расходятся к полюсам, образуя гаметы с числом хромосом от 0 до 18. Большинство их нежизнеспособно. Лишь изредка формируются мужские и женские гаметы, имеющие сбалансированное нередуцированное число хромосом 9R+9B. При их слиянии в процессе оплодотворения образуется зигота, из которой развивается аллотетраплоид с генотипом ♀(9R+9B) +, ♂(9R+ 9В). При созревании его половых клеток в мейозе каждая хромосома из R генома отца будет коньюгировать с гомологичной ей хромосомой R генома матери, а гомологичные хромосомы генотипов В отца и матери будут коньюгировать между собой. Процесс мейоза завершается нормально с образованием гамет с генотипом 9R +9В. Плодовитость редечно-капустного гибрида окажется восстановленной. Впервые межродовой плодовитый аллополиллоид при скрещивании редьки и капусты был получен Г.Д.Карпеченко в 1928 г. Он получил первый искусственным полиплоид.
Полиплойдные организмы, как правило, обладают повышенной жизнеспособностью, более крупными размерами, ус- тойчивостью к неблагоприятным факторам среды. Их генотипы обуславливают более широкую норму реакции, а при поло- вом размножении нередко наблюдается высокая степень изменчивости с образованием большого числа генотипических и фенотипических классов. Поэтому полиплоидия имеет большое значение в эволюции дикорастущих и культурных растений.
Около половины покрытосеменных растений являются полиплоидами. В северных широтах и высокогорных районах полиплоиды составляют 80% произрастающих там растений.
Большинство культурных растений также полиплойды. К ним относятся, например, пшеница, кукуруза, сахарная свекла, картофель, плодовые культуры и др. Декоративные цветковые растения с очень крупными цветками: тюльпаны, нарциссы, гладиолусы и др. - также полиплоиды.
Виды многих родов культурных растений представляют собой полиплоидные ряды. Например, наиболее, примитивная диплоидная форма культурных пшениц имеет 14 хромосом, твердая пшеница содержит 28 хромосом (тетраплоид), а мягкая -42 хромосомы (гексагоюид). Основное гаплоидное число хромосом в роде пшениц равно 7.
У животных полиплоидия встречается редко. В основном она наблюдается среди гермафродитов (например, у земляных червей) и у видов, самки которых размножаются путем партеногенеза (некоторые жуки, бабочки, клопы, ракообразные, рыбы и саламандры).
-
Причинами, затрудняющими распространение полиплоидии у животных служат:
-
нарушение баланса между половыми хромосомами и аутосомами, необходимого для определения пола;
-
перекрестное оплодотворение у большинства животных, приводящее к нарушению конъюгации хромосом;
-
более сложный процесс онтогенеза, который при полиплоидии чаще нарушается;
-
нежизнеспособность или стерильность гибридов животных.
Анеуплоидия (от греч. an - отрицательная частица, ей - вполне, pllos - кратный и eidos - вид), гетероплоидия (от греч. heteros - иной, другой; означает разнородность) - геномная мутация, состоящая в изменении числа хромосом, некратным гаплойдному. В результате этой мутации в хромосомном наборе одна или несколько хромосом отсутствуют или, наоборот, имеются в избытке.
Таким образом, гетероплоидия обусловлена изменением в геноме количества отдельных хромосом. При гетероплои- дии в нормальном хромосомном наборе либо отсутствуют или имеются в избыточном количестве одна или более хромосом. Различают моносомию (2n-1), нуклисомию (2n-2), трисомию (2n+1) и полисомию (2n+х) по отдельным хромосомам. Утрата одной хромосомы в диплоидном наборе (2n-1) называется моносомией, а организм - моносомиком. Причиной указанных геномных мутаций служит нарушение расхождения хромосом в мейозе. При отсутствии двух негомологичных хромосом (2п-2) организм является двойным моносомиком, а при отсутствии пары гомологичных хромосом - нуллисомиком. Наличие в наборе трех гомологичных хромосом называется трисомией, а организм - трисомиком. Трисомия по двум негомологичным хромосомам (2п+2) наблюдается у двойных трисомиков. Тройные трисомики имеют генотип (2п+3). Термины тетрасрмик, пентасомик, полисомик означают, что в хромосомном наборе присутствуют соответственно, 4, 5 или большее число дащних хромосом.
При анеууплоидии в нормальном хромосомном наборе либо отсутствует одна или более хромосом, либо присутствует одна или более добавочных хромосом. Организмы, у которых отсутствует одна пара хромосом называют "нуллисомиками", и “моносомиками”, если отсутствует одна хромосома. Таким образом, понятия трисомик, тетрасомик и т.д. означают, что э хромосомном наборе присутствует соответственно одна, две и т.д. лишние хромосомы.
Ануеплоиды могут возникнуть, если в анафазе I мейоза гомологичные хромосомы одной или нескольких пар не разойдутся и вместе направятся к одному из полюсов клетки. Например, нерасхождение какой-то пары гомологичных хромо* сом может привести к появлению гаметы, лишенной данной хромосомы, и другой гаметы, имеющей эту-хромосому в двойном количестве. Аналогичные изменения могут наблюдаться при нерасхождении сестринских хроматид в анафазе второго мейотического деления. В результате образуются гаметы с избыточным или недостаточным числом хромосом. Когда такие гаметы сливаются с нормальными гаметами, образуются зиготы с нечетным числом хромосом. Таким образом, при оплодотворении таких гамет нормальными половыми клетками образуются зиготы, в кариотипе которых изменено общее число хромосом за счет уменьшения (моносомия) или увеличения (трисомия) числа отдельных хромосом.
Геномные мутации изменяют баланс генов и тем самым нарушают процесс индивидуального развития организма. Большинство зигот с недостаточным числом хромосом обычно нежизнеспособны. Зиготы с лишними хромосомами иногда способны к развитию. Но развивающиеся из них анеуплоиды характеризуются пониженной жизнеспособностью и рядом резко выраженных аномалий. У человека такие мутации (приводят к анеуплоидии) являются причиной возникновения многих тяжелых болезней (синдром Дауна, синдром Эдварса, синдром Патау и др.)