1
.docx|
1)Качественные особенности живой материи 1. Химический состав. Живые существа состоят из тех же химических элементов, что и неживые, но в организмах есть молекулы веществ, характерных только для живого (нуклеиновые кислоты, белки, липиды) . 2. Дискретность и целостность. Любая биологическая система (клетка, организм, вид и т. д. ) состоит из отдельных частей, т. е. дискретна. Взаимодействие этих частей образует целостную систему (например, в состав организма входят отдельные органы, связанные структурно и функционально в единое целое) . 3. Структурная организация. Живые системы способны создавать порядок из хаотичного движения молекул, образуя определенные структуры. Для живого характерна упорядоченность в пространстве и времени. Это комплекс сложных саморегулирующихся процессов обмена веществ, протекающих в строго определенном порядке, направленном на поддержание постоянства внутренней среды — гомеостаза. 4. Обмен веществ и энергии. Живые организмы — открытые системы, совершающие постоянный обмен веществом и энергией с окружающей средой. При изменении условий среды происходит саморегуляция жизненных процессов по принципу обратной связи, направленная на восстановление постоянства внутренней среды — гомеостаза. Например, продукты жизнедеятельности могут оказывать сильное и строго специфическое тормозящее воздействие на те ферменты, которые составили начальное звено в длинной цепи реакций. 5. Самовоспроизведение. Самообновление. Время существования любой биологической системы ограничено. Для поддержания жизни происходит процесс самовоспроизведения, связанный с образованием новых молекул и структур, несущих генетическую информацию, находящуюся в молекулах ДНК. 6. Наследственность. Молекула ДНК способна хранить, передавать наследственную информацию, благодаря матричному принципу репликации, обеспечивая материальную преемственность между поколениями. 7. Изменчивость. При передаче наследственной информации иногда возникают различные отклонения, приводящие к изменению признаков и свойств у потомков. Если эти изменения благоприятствуют жизни, они могут закрепиться отбором. 8. Рост и развитие. Организмы наследуют определенную генетическую информацию о возможности развития тех или иных признаков. Реализация информации происходит во время индивидуального развития — онтогенеза. На определенном этапе онтогенеза осуществляется рост организма, связанный с репродукцией молекул, клеток и других биологических структур. Рост сопровождается развитием. 9. Раздражимость и движение. Все живое избирательно реагирует на внешние воздействия специфическими реакциями благодаря свойству раздражимости. Организмы отвечают на воздействие движением. Проявление формы движения зависит от структуры организма. |
6)Концепции происхождения жизни теория, построенная на целом ряде экспериментальных исследований ученых разных стран, в основе которой лежит гипотеза акад. А. И. Опарина о возникновении жизни на Земле абиогенным путем. В опытах ученых Дж. Холдейна, С. Фокса, С. Миллера, Г. Мёллера удалось воспроизвести физико-химические условия, в которых из неорганических веществ возникали органические, образовывавшие, в свою очередь, подобие коацерватов.(белки ,жиры ,углеводы ,нуклеиновые кислоты концентрируются-образуют коацерваты) Основные положения теории следующие: 1) органические вещества сформировались из неорганических под действием физических факторов среды; 2) органические вещества взаимодействовали, образуя все более сложные вещества, в результате чего возникли ферменты и самовоспроизводящиеся системы — свободные гены; 3) свободные гены соединялись с другими высокомолекулярными органическими веществами; 4) вокруг них стали образовываться белково-липидные мембраны; 5) возникли клетки Первый этап возникновения жизни на Земле согласно гипотезе А. И. Опарина, это образование органических веществ из неорганических, происходившее в водах первичного океана более 3,5 млрд. лет назад. При этом на Землю, которую еще не защищал озоновый экран, свободно проникало ультрафиолетовое излучение; в атмосфере происходили грозовые разряды. В условиях бескислородной среды атмосфера насыщалась альдегидами, спиртами, аминокислотами. Второй этап возникновения жизни на земле -согласно гипотезе А.И. Опарина-этап образования из простых органических соединений в водах первичного океана белков ,жиров , углеводов, нуклеиновых кислот. Разрозненные молекулы соединений концентрировались и формировали коацерваты, действующие как открытые системы, способные к обмену веществ с окружающей средой и к росту. Третий этап возникновения жизни на Земле согласно гипотезе А. И. Опарина, этап развития жизни, когда коацерваты соединялись, укрупнялись, взаимодействовали между собой и с другими веществами, поглощаемыми ими. В результате взаимодействия коацерватов с нуклеиновыми кислотами образовались первые живые существа — пробионты, способные, помимо роста и обмена веществ, к самовоспроизведению. Наступил период органической эволюции, в основе которой лежали изменчивость, наследственность, естественный отбор. В отложениях конца архейской эры (2,7-3,5 млрд. лет назад) находят следы предъядерных организмов (синезеленые. хемотрофные и гетеротрофные бактерии). В результате прогрессивных усложнений появились автотрофные растительные организмы, а вместе с ними свободный кислород и органические вещества — продукты фотосинтеза. Организмы, оставшиеся гетеротрофными, дали начало грибами и животным.
|
32)Хлоропласты как органеллы трансформации световой энергии в в энергию химических связей Процесс фотосинтеза осуществляется в хлоропластах в два этапа. В гранах (тилакоидах) протекают реакции, вызываемые светом.- световые, а в строме - реакции, не связанные со светом,- темновые, или реакции фиксации углерода. Световые реакции. 1. Свет, попадая на молекулы хлорофилла, которые находятся в мембранах тилакоидов гран, приводит их в возбужденное состояние. В результате этого электроны е сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где и накапливаются, создавая отрицательно заряженное электрическое поле. 2. Место вышедших электронов в молекулах хлорофилла занимают электроны воды е, так как вода под действием света подвергается фоторазложению (фотолизу): Гидроксилы ОН~, став радикалами ОН, объединяются: 4ОН--> 2H20+O2, образуя воду и свободный кислород, который выделяется в атмосферу. 3. Протоны водорода H+ не проникают через мембрану тилакоида и накапливаются внутри, образуя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны. 4. При достижении критической разности потенциалов протоны Н"^ устремляются по протонному каналу в ферменте АТФ-синтетаза, встроенному в мембрану тилакоида, наружу. На выходе из протонного канала создается высокий уровень энергии, которая идет на синтез АТФ (АДФ+Ф АТФ). Образовавшиеся молекулы АТФ переходят в строму, где участвуют в реакциях фиксации углерода. 5. Протоны Н"^, вышедшие на поверхность мембраны тилакоида, соединяются с электронами в, образуя атомарный водород Н, который идет на восстановление переносчика НАДФ+, 2e+H+ + HAДФ+ ---> HAДФ•H (переносчик с присоединенным водородом) Таким образом, активированный световой энергией электрон хлорофилла используется для присоединения водорода к переносчику. HAДФ•H переходит в строму хлоропласта, где участвует в реакциях фиксации углерода. Реакции фиксации углерода (темновые реакции). Осуществляются в строме хлоропласта, куда поступают АТФ, НАДФ-Н от тилакоидов гран и СОз из воздуха. Кроме того, там постоянно находятся пятиуглеродные соединения - пентозы C5, которые образуются в цикле Кальвина (цикле фиксации СO2). Этот цикл можно проследить на углероде как главном элементе углеводов. 1. К пентозе С5 присоединяется СO2. в результате чего появляется нестойкое шестиуглеродное соединение С6, которое расщепляется на две трехуглеродные группы 2Сз - триозы. 2. Каждая из триоз 2Сз принимает по одной фосфатной группе от 2 АТФ, что обогащает молекулы энергией. 3. Каждая из триоз 2Сз присоединяет по одному атому водорода от 2 НАДФ-Н. 4. После чего одни триозы объединяются, образуя углеводы (2Сз --> С6 --> С6Н120б) (глюкоза) 5. Другие триозы объединяются, образуя пентозы 5Сз --> 3С5, и вновь включаются в цикл фиксации СО2. Суммарная реакция фотосинтеза: Хемосинтез. Некоторые бактерии, лишенные хлорофилла, тоже способны к синтезу органических соединений, при этом они используют энергию химических реакций неорганических веществ. Преобразование энергии химических реакций в химическую энергию синтезируемых органических соединений называется хемосинтезом.
|
||
|
2) Уровни организации жизни. Уровни организации живых систем представляют собой некую упорядоченность, иерархическую систему, которая является одним из основных свойств живого. Уровни жизни отличаются друг от друга сложностью организации системы. Уровень жизни – это форма и способ ее существования. Например, вирус существует в виде молекулы ДНК или РНК, заключенной в белковую оболочку. Это форма существования вируса. 1)Молекулярно-генетический уровень представлен отдельными биополимерами (ДНК, РНК, белками, липидами, углеводами и другими соединениями); на этом уровне жизни изучаются явления, связанные с изменениями (мутациями) и воспроизведением генетического материала, обменом веществ. 2)Клеточный – уровень, на котором жизнь существует в форме клетки – структурной и функциональной единицы жизни. На этом уровне изучаются такие процессы, как обмен веществ и энергии, обмен информацией, размножение, фотосинтез, передача нервного импульса и многие другие. 3)Тканевый уровень – совокупность клеток с одинаковым типом организации составляет ткань. Тканевый уровень возник вместе с появлением многоклеточных животных и растений, имеющих различающиеся между собой ткани. Большое сходство между всеми организмами сохраняется на тканевом уровне. 4)Органный уровень – совместно функционирующие клетки, относящиеся к разным тканям, составляют органы. 5)Организменный – это самостоятельное существование отдельной особи – одноклеточного или многоклеточного организма. 6)Популяционно-видовой – уровень, который представлен группой особей одного вида – популяцией; именно в популяции происходят элементарные эволюционные процессы – накопление, проявление и отбор мутаций. 7)Биогеоценотический – представлен экосистемами, состоящими из разных популяций и среды их обитания. 8)Биосферный – уровень, представляющий совокупность всех биогеоценозов. В биосфере происходит круговорот веществ и превращение энергии с участием организмов. Продукты жизнедеятельности организмов участвуют в процессе эволюции Земли. |
3)Современное представление о эукариотах и прокариотах, их генезисе. Структурно каждая живая клетка состоит, как минимум, из трех частей: ядра, цитоплазмы (внутренней среда клетки) и ее оболочки (или мембраны). Если ядро клетки ярко не выражено, ДНК практически равномерно заполняет весь внутренний объем клетки (первая стадия клеточной жизни), а сами клетки практически ничем не отличаются друг от друга, организмы называются прокариотами. При большей выраженности клеточных ядер и наличии в них нескольких хромосом (ДНК с окружающими их РНК и белками), организмы называются эукариотами. Под влиянием внешних условий некоторые признаки отдельных ДНК в одной или нескольких хромосомах могут немного изменяться. В конце концов, эти различия приводят к образованию организмов одного биологического вида, но различающихся по полу. У эукариотов большая гибкость клеточных оболочек, а, значит, - и их подвижность (поэтому большинство из них - все-таки, животные). Эта гибкость и податливость доходит до того, что некоторые из них способны не только всасывать простые вещества и органические молекулы, но и поглощать целые другие клетки меньшего размера. (Они и стали первыми хищниками). Из двух последних подразделов вытекает, что все хищники - гетеротрофы и эукариоты. Эукариоты есть и среди растительного, и среди животного мира, хотя среди одноклеточных между теми и другими бывает очень трудно провести четкую границу. Типичным пороговым (находящимся между растениями и животными) одноклеточным организмом является эвглена зеленая .Она, как растение, содержит хлорофилл и при дневном свете способна к фотосинтезу, что определяет ее автотрофный тип питания в этих условиях. Однако, в темноте эвглена питается только осмотическим путем (всасывая полезные вещества из окружающей среды через оболочку клетки). Это относит ее к организмам с гетеротрофным типом питания (обычно такие организмы бывают животными).
|
5)Критерии жизни
Жизнь – это макромолекулярная открытая система, которой свойственна иерархическая организация, способность к самовозобновлению, обмен веществ и тонко регуляторный процесс. Критерии жизни 1. Самовозобновление, которое связано с постоянным обменом вещества и энергии, и в основе которого лежит способность хранить и использовать биологическую информацию в виде уникальных информационных молекул: белков и нуклеиновых кислот. 2. Самовоспроизведение, которое обеспечивает преемственность между поколениями биологических систем 3. Саморегуляция, которая основана на потоке вещества, энергии и информации 4. Большинство химических процессов в организме находятся не в динамичном состоянии 5. Живые организмы способны к росту 6. Обмен веществом и энергией 7. Обмен веществ – особый способ взаимодействия живых организмов со средой 8. Обмен веществ требует постоянного притока некоторых веществ и энергии из вне и выделения некоторых продуктов диссимиляции во внешнюю среду. Организм является открытой системой 9. Раздражимость – заключается в передаче информации от внешней среды к организму; на основе раздражимости осуществляется Саморегуляция и гомеостаз 10. Репродукция – воспроизведение себе подобных 11. Наследственность – поток информации между поколениями в результате чего обеспечивается преемственность 12. Изменчивость – появление новых признаков в процессе репродукции; основа эволюции 13. Онтогенез – индивидуальное развитие, реализация индивидуальной программы 14. Филогенез – историческое развитие, эволюционное развитие осуществляется в результате наследственной изменчивости, естественного отбора и борьбы за существование 15. Организмы включены в процесс эволюции
|
||
|
17)Цикл кребса, его значение Ци́кл трикарбо́новых кисло́т (цикл Кре́бса, цитра́тный цикл) — центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии — АТФ. Цикл Кребса — это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др. |
19)Акцепторы водорода( в митохондриях) Формулы акцепторов водорода: НАД, ФАД, КоQ. Роль кислорода в цепи переноса электронов. Дыхательная цепь – последовательная цепь ферментов, осуществляющая перенос ионов водорода и электронов от окисляемых субстратов к молекулярному кислороду – конечному акцептору водорода. В ходе этих реакций выделение энергии происходит постепенно, небольшими порциями, и она может быть аккумулирована в форме АТФ. Локализация ферментов дыхательной цепи – внутренняя митохондриальная мембрана. I. НАДН-KoQ-редуктаза (содержит промежуточные акцепторы водорода: флавинмононуклеотид и железосерные белки). II. Сукцинат-KoQ-редуктаза (содержит промежуточные акцепторы водорода: ФАД и железосерные белки). III. KoQН2-цитохром с-редуктаза (содержит акцепторы электронов: цитохромы b и с1, железосерные белки). IV. Цитохром с-оксидаза (содержит акцепторы электронов: цитохромы а и а3, ионы меди Cu2+).
|
13)Цикличность превращения АТФ в АДФ и АДФ в АТФ. При отделении одног остатка фосфорной кислоты АТФ переходит в АДФ(,если отедляется еще один остаток фосфорной кислоты АДФ переходит в АМФ,что бывает крайне редко) Место отделившегося остатка фосфорной кислоты занимает молекула воды. Отделение каждого остатка фосфорной кислоты происходит с помощью ферментов, при этом выделяется 40 кДж( а при разрыве обычных ковалентных связей – около 12 кДж энергии).При синтезе АТФ,наоборот, поглощается большое количество энергии. У всех организмов АТФ синтезируется на внутренней мембране митохондрий в процессе кислородного(III)этапа диссимиляции(катаболизма),поэтому их называют энергетическими станциями клетки. (2С3Н4О3+6О2+36АДФ+36Ф6СО2+36АТФ+42Н2О) |
||
|
7)Биосинтез белка. Основные этапы:транскрипция,пронессии( созревание и-РНК) Биосинтез белка – сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК. Именно туда поступает информационная РНК из ядра клетки. Биосинтез белка можно разделить на стадии транскрипции, процессинга и трансляции. Транскрипция (от лат. transcriptio — переписывание) — биосинтез молекул РНК ,осуществляемый на молекулах ДНК по принципу матричного синтеза. При помощи ферментов на соответствующих участках молекулы ДНК(генах) синтезируются все виды РНК. Синтезируется 20 разновидностей тРНК,так как в биосинтезе белка принимают участие 20 аминокислот. Затем иРНК и тРНК выходят в цитоплазму, рРНК встраивается в субъединицы рибосом, которые также выходят в цитоплазму. Обратная транскрипция — это процесс образования двуцепочечной ДНК на матрице одноцепочечной РНК. Данный процесс называется обратной транскрипцией, так передача генетической информации при этом происходит в «обратном» направлении, относительно транскрипции. Процессинг (англ. processing — обработка, переработка)— процесс формирования зрелых молекул РНК из их предшественников (пре-РНК). Иными словами, это совокупность реакций, ведущих к превращению первичных продуктов транскрипции (т. е. пре-РНК различных видов) в функционирующие молекулы. Процессинг т- и рРНК в основном сводится к удалению лишних фрагментов с концов молекул. Что касается иРНК, то у эукариот ее процессинг осуществляется многоступенчато. Основными его событиями являются следующие: — модификация концов молекулы и РНК, в ходе которой к концам молекулы присоединяются специфические короткие, последовательности нуклеотидов, обозначающие место начала и место конца трансляции; - сплайсинг - удаление неинформативных последовательностей РНК, соответствующих интронам ДНК. У прокариот иРНК не подвергаются процессингу — они способны работать сразу после синтеза. У всех организмов процессинг РНК происходит в ядре. Для каждого типа молекул он осуществляется специальным ферментом (или группой ферментов) (Процессингу также могут подвергаться и продукты трансляции, т . е. полипептиды, непосредственно считанные с иРНК. Таким изменениям подвергаются молекулы — предшественники многих белков - пищеварительных ферментов, коллагена, некоторых гормонов, иммуноглобулинов и др., после чего они начинают реально функционировать в организме). |
8)Биосинтез белка:трансляция, предрибосомный и рибосомный период. Биосинтез белка – сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК. Именно туда поступает информационная РНК из ядра клетки. Биосинтез белка можно разделить на стадии транскрипции, процессинга и трансляции. Транскрипция – это процесс снятия информации с молекулы ДНК, синтезируемой на ней молекулой и-РНК. Информационная РНК состоит из одной цепи и синтезируется на ДНК в соответствии с правилом комплементарности. Как и в любой другой биохимической реакции в этом синтезе участвует фермент. Он активирует начало и конец синтеза молекулы и-РНК. Готовая молекула и-РНК выходит в цитоплазму на рибосомы, где происходит синтез полипептидных цепей. Процесс перевода информации, содержащейся в последовательности нуклеотидов и-РНК, в последовательность аминокислот в полипептиде называется трансляцией. Аминокислоты доставляются к рибосомам транспортными РНК. Эти РНК имеют форму клеверного листа. На конце молекулы есть площадка для прикрепления аминокислоты, а на вершине – триплет нуклеотидов, комплементарный определенному триплету – кодону на и-РНК. Этот триплет называется антикодоном. Ведь он расшифровывает код и-РНК. В клетке т-РНК всегда столько же, сколько кодонов, шифрующих аминокислоты. Рибосома движется вдоль и-РНК, смещаясь при подходе новой аминокислоты на три нуклеотида, освобождая их для нового антикодона. Аминокислоты, доставленные на рибосомы, ориентированы по отношению друг к другу так, что карбоксильная группа одной аминокислоты оказывается рядом с аминогруппой другой аминокислоты. В результате между ними образуется пептидная связь. Постепенно формируется молекула полипептида. Синтез белка продолжается до тех пор, пока на рибосоме не окажется один из трех стоп-кодонов – УАА, УАГ, или УГА. После этого полипептид покидает рибосому и направляется в цитоплазму. На одной молекуле и-РНК находятся несколько рибосом, образующих полисому. Именно на полисомах и происходит одновременный синтез нескольких одинаковых полипептидных цепей. Каждый этап биосинтеза катализируется соответствующим ферментом и обеспечивается энергией АТФ. Биосинтез происходит в клетках с огромной скоростью. В организме высших животных в одну минуту образуется до 60 тыс. пептидных связей |
68)Полимерия Полимерия — явление, когда различные неаллельные гены могут оказывать однозначное действие на один и тот же признак, усиливая его проявление. Наследование признаков при полимерном взаимодействии генов. В том случае, когда сложный признак определяется несколькими парами генов в генотипе и их взаимодействие сводится к накоплению эффекта действия определенных аллелей этих генов, в потомстве гетерозигот наблюдается разная степень выраженности признака, зависящая от суммарной дозы соответствующих аллелей. Например, степень пигментации кожи у человека, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4) (см. рис. 3.80). При браке двух мулатов, гетерозиготных по всем четырем парам, которые образуют по 24 = 16 типов гамет, получается потомство, 1/256 которого имеет максимальную пигментацию кожи, 1/256 — минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака. В разобранном примере доминантные аллели полигенов определяют синтез пигмента, а рецессивные — практически не обеспечивают этого признака. В клетках кожи организмов, гомозиготных по рецессивным аллелям всех генов, содержится минимальное количество гранул пигмента. В некоторых случаях доминантные и рецессивные аллели полигенов могут обеспечивать развитие разных вариантов признаков. Например, у растения пастушьей сумки два гена одинаково влияют на определение формы стручочка. Их доминантные аллели образуют одну, а рецессивные — другую форму стручочков. При скрещивании двух дигетерозигот по этим генам (рис. 6.16) в потомстве наблюдается расщепление 15:1, где 15/16 потомков имеют от 1 до 4 доминантных аллелей, а 1/16, не имеет доминантных аллелей в генотипе. Если гены располагаются, каждый в своем отдельном локусе, но их взаимодействие проявляется в одном и том же направлении – это полигены. Один ген проявляет признак незначительно. Полигены дополняют друг друга и оказывают мощное действие – возникает полигенная система – т.е. система является результатом действия одинаково направленных генов. Гены подвергаются значительному влиянию главных генов, которых более 50. полигенных систем известно множество. При сахарном диабете наблюдается умственная отсталость. Рост, уровень интеллекта - определяются полигенными системами |
||
|
15)Брожение Броже́ние — метаболический процесс, при котором регенерируется АТФ, а продукты расщепления органического субстрата могут служить одновременно и донорами, и акцепторами водород. Брожение — это анаэробный (происходящий без участия кислорода) метаболический распад молекул питательных веществ, например глюкозы. По выражению Луи Пастера, «брожение — это жизнь без кислорода». Большинство типов брожения осуществляют микроорганизмы — облигатные или факультативные анаэробы. Броже́ние — процесс анаэробного расщепления органических веществ, преимущественно углеводов, происходящий под влиянием микроорганизмов или выделенных из них ферментов. Биохимический процесс, вызываемый микроорганизмами и приводящий к распадению органических веществ. Спиртовое брожения уравнение C6H12O6+2H3PO4+2AДФ=2C2H5OH+2CO2+2АТФ+2H2O
|
11)Энергодонорные системы клетки Митохо́ндрия— двумембранный сферический или эллипсоидный органоид диаметром обычно около 1 микрон. Характерна для большинства эукариотических клеток, как автотрофов (фотосинтезирующие растения), так и гетеротрофов (грибы, животные). Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза АТФ и термогенеза. Эти три процесса осуществляются за счёт движения электронов по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается[1]: так, одноклеточные зелёные водоросли (эвглена, хлорелла, политомелла) и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300 000 и 500 000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. В специализированных клетках органов животных содержатся сотни и даже тысячи митохондрий (мозг, сердце, мышцы). |
22)Фотосинтез как процесс планетарного значения Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф) , также является запасённой в процессе фотосинтеза. Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу. |
||
|
10)Регуляция биосинтеза ,ее уровни. Антибиотики: их роль в регуляции синтеза белка патогенных микроорганизмов Белки(полипептиды) 1(протеины) Биополимеры,мономеры являются аминокислоты +(-) 200 аминокислот 20 входят в состав белков-протеинногенные Н (амино)Н2N-С-СООН(карбоксильная) R Протеиногенные аминокислоты: 1-аланин-а/а 2-валин-val 3-глицин-gly 4-лицин-lys 5-лейцин-leu 6-пролин-pro 7-аргинин-arg 8-треонин-thr 9-триптофан-trp 10-серин-ser 11-метионин-met 12-цистеин-cys 13-фенилаланин-phe 14-аспарагин-asn 15-аспартат-asp 16-глутамин-gln 17-глутамат-glu 18-гистидин-hiss 19-гуолейцин-ile 20-гидроксипролин-hyp Уровни организации белков: 1)первичная(линейная цепочка,пептидная связь) 2)вторичная(конфериация-пространственная укладка белков) 3)третичная(трехмерная укладка) 4)четвертичная Два типа вторичной связи: 1)a(альфа)-спираль 2)B(бета)-складчатость |
9)т-РНК как адаптор и акецептор Молекулы т-РНК являются молекулами адапторами, т.е. посредниками, при помощи которых АК переносятся и включаются в определенном порядке в растущую полипептидную цепь. Транспортная РНК - молекула-адаптор, или дешифровщик. По¬скольку между нуклеотидами и аминокислотами невозможны специфиче¬ские взаимодействия (по типу комплементарных нуклеотидных пар), то появилось предположение о существовании молекул-адапторов, которые могут взаимодействовать как с определенным кодоном, так и с определен¬ной аминокислотой. Такими молекулами оказались транспортные РНК. 3'-Конец всех их молекул заканчивается одинаковой последовательностью ЦЦА, к которой могут присоединяться аминокислоты. Каждая т-РНК специфично связыва¬ется только с одной определенной аминокислотой, а общее количество ви¬дов т-РНК - 61, т.е. столько же, сколько имеется смысловых кодонов. В то же время одна аминокислота может взаимодействовать с двумя-шестью разными т-РНК, что объясняет вырожденность кода. Каждая тРНК присоединяет определённую аминокислоту и транспортирует её к месту сборки полипептида в рибосоме. В молекуле тРНК есть два активных участка: триплет-антикодон на одном конце и акцепторный конец на другом. Антикодон считывает информацию с иРНК, акцепторный конец является посадочной площадкой для аминокислоты. Синтез полипептидной цепи белковой молекулы начинается с активации аминокислот, которую осуществляют специальные ферменты. Каждой аминокислоте соответствует как минимум один фермент. Фермент обеспечивает присоединение аминокислоты к акцепторному участку тРНК с затратой энергии АТФ. |
21)Регуляция синтеза АТФ Работа дыхательных ферментов регулируется с помощью эффекта, который получил название дыхательный контроль. Дыхательный контроль – это прямое влияние электрохимического градиента на скорость движения электронов по дыхательной цепи (т.е. на величину дыхания). В свою очередь, величина градиента напрямую зависит от соотношения АТФ и АДФ, количественная сумма которых в клетке практически постоянна ([АТФ] + [АДФ] = const). Реакции катаболизма направлены на поддержание постоянно высокого уровня АТФ и низкого АДФ. Возрастание протонного градиента возникает при снижении количества АДФ и накоплении АТФ (состояние покоя), т.е. когда АТФ-синтаза лишена своего субстрата и ионы Н+ не проникают в матрикс митохондрии. При этом ингибирующее влияние градиента усиливается и продвижение электронов по цепи замедляется. Ферментные комплексы остаются в восстановленном состоянии. Следствием является уменьшение окисления НАДН и ФАДН2 на I и II комплексах и замедление катаболизма в клетке. Снижение протонного градиента возникает при исчерпании резервов АТФ и избытке АДФ , т.е. при работе клетки. В этом случае активно работает АТФ-синтаза и через канал Fо проходят в матрикс ионы Н+. При этом градиент, естественно, снижается, поток электронов возрастает, в результате повышается выкачивание ионов Н+ в межмембранное пространство и снова их быстрое "проваливание" через АТФ-синтазу внутрь митохондрий с синтезом АТФ. Ферментные комплексы I и II усиливают окисление НАДН и ФАДН2 (как источников электронов) и снимается ингибирующее влияние НАДН на цикл лимонной кислоты и пируватдегидрогеназный комплекс. Как итог – активируются реакции катаболизма углеводов и жиров.
|
||
|
57) Цикл развития свиного цепня, пути зараженияхозяев. Лабораторная диагностика. Цистеркоз(цистистеркоз). Профилактика заболеваний Свиной цепень (Taenia solium) во взрослом состоянии паразитирует в тонком кишечнике человека и вызывает заболевание - тениоз. Длина его достигает 3 - 7 метров. На головке располагаются 4 присоски и венчик крючьев. За головкой находится шейка, за шейкой тело, состоящее из члеников. Системы органов такие же, как у бычьего цепня. Но в отличие от бычьего цепня, в гермафродитных члениках свиного цепня три дольки яичника, а зрелый членик свиного цепня имеет 7 - 12 ответвлений матки. В зрелых члениках формируются яйца с шестикрючным зародышем - онкосферой. Для дальнейшего развития личинка должна попасть в промежуточного хозяина - свинью. В желудке свиньи оболочки яиц растворяются, личинки с током крови попадают в мышцы и превращаются в финну - цистицерк. Человек заражается личинками свиного цепня, если съедает непроваренное мясо свиньи. Иногда во время рвоты из кишечника человека возможно забрасывание отдельных члеников свиного цепня в желудок, где они перевариваются. Оболочки яиц растворяются, личинки выходят из них и с током крови попадают в различные органы (мозг, печень, легкие, глаза), где формируются финны - цистицерки, вызывая заболевание - цистицеркоз. Заражения свиным цепнем можно избежать, если не употреблять непроваренное мясо свиньи.
|
29)Хемосинтез: реакции,связанные с железо- и серобактерияими Имеются также бактерии, способные окислять двухвалентное железо до трехвалентного. Общее уравнение хемосинтеза в этом случае выглядит так: 4FeCO3+O2+6H2O=4Fe(OH)3+4CO2+324кДж Такие бактерии, живущие в болотах, образуют болотные месторождения железа В результате деятельности железобактерий образуется Fe(OH)3, скопления которого образуют болотную железную руду. 2H2S + 02 =2H20 + 2S + 272 кДж Бесцветные серобактерии окисляют сероводород и накапливают в своих кллетках сперуу. Выделяющаяся в результате свободная сера накапливается в их клетках в виде множества крупинок. При недостатке сероводорода бесцветные серобактерии производят дальнейшее окисление находящейся в них свободной серы до серной кислоты. При недостатке сероводорода бактерии производят дальнейшее окисление серы до серной кислоты 2S+302+2H20 =2H2S04 + 636 к Дж Образовавшаяся в результате энергия также используется на синтез органического вещества из углекислого газа. Энергетический эффект окисления сероводорода до серной кислоты равен 666 кДж на каждую окисленную грамм-молекулу сероводорода. Колоссальное количество серобактерий имеется в Черном море , в котором глубже 200 м (а в некоторых местах почти от поверхности) вода насыщена сероводородом. Серобактерии,образуя серную кислоту,способствуют постепенному разрушению и выветриванию горных пород, разрушению каменных и металлических сооружений, окисляющие серу до сульфатов,участвуют в очищении промышленных сточных вод.
|
38)Гаметогенез: образование мужских гамет . Оплодотворение. Половой диморфизм и биологическое значение разнополости. Оплодотворение – это процесс слияния половых клеток. Процесс оплодотворения складывается из трех последовательных фаз: сближения гамет, активации яйцеклетки, слияния гамет или сингамии. Случайная встреча разных гамет при оплодотворении приводит к тому, что среди особей вида практически невозможно появление двух генотипически одинаковых организмов. Достигаемое с помощью описанных процессов генотипическое разнообразие особей предполагает наследственные различия между ними на базе общего видового генома. Половой диморфизм -(греч. «ди» —два, «морфе» форма)- - наличие у одного и того же вида животных резких различий по величине, окраске, внешнему виду между особями мужского и женского пола. Половой диморфизм особенно выражен у павлинов, фазанов, уток. У этих птиц самцы значительно крупнее и красивее самок. У львов гриву имеют только самцы. У певчих птиц поют только самцы. Явные признаки, указывающие на пол, называют вторичными половыми признаками. Эти признаки хорошо выражены и у человека. Ч. Дарвин изучил половой диморфизм на примере птиц и бабочек. Он рассматривал его как следствие полового отбора. Биологическое значение разнополости: 1)возможность продолжения рода 2)Возможность создания более жизнеспособного потомства, благодаря гемизиготности.(хх,ху) 3)обеспечивает возможность полового размножения
|
||
|
23)Энергетическая фаза фотосинтеза Световая фаза фотосинтеза осуществляется в хлоропластах, где на мембранах расположены молекулы хлорофилла. Хлорофилл поглощает энергию солнечного света. Эта энергия используется на синтез молекул АТФ из АДФ и фосфорной кислоты и способствуют расщеплению молекул воды: 2H20=4H++4+O2. Образующийся при этом кислород выделяется в окружающую среду.
Под влиянием энергии солнечного света молекула хлорофилла возбуждается, и один из ее электронов переходит на более высокий энергетический урввень. Богатый энергией электрон участвует в окислительно-восстановительных реакциях и отдает избыточную энергию, проходя по цепи переносчиков электронов. Эта цепь образована различными белками, встроенными во внутреннюю мембрану хлоропласта. Отдаваемая электроном энергия используется на синтез молекул АТФ. Таким образом, энергия солнечного света необходима для перемещения электронов по цепи переносчиков электронов. При этом световая энергия преобразуется в химическую и запасается в молекулах АТФ. Молекулы хлорофилла, потерявшие электроны, присоединяют электроны, образующиеся при расщеплении молекулы воды. Процесс расщепления молекул воды под влиянием солнечной энергии называют фотолизом (от греч. fotos - свет и lisis - растворение). В результате фотолиза образуются: Электроны, заполняющие "дырки" в молекулах хлорофилла. Протоны H+, которые соединяются с веществом НАДФ+ - переносчиком ионов водорода и электронов и восстанавливают его до НАДФ•Н. Молекулярный кислород, который выделяется в окружающую среду. Таким образом, в результате световой фазы фотосинтеза восстанавливается НАДФ+ и образуется НАДФ•Н, синтезируется АТФ из АДФ и фосфорной кислоты, выделяется молекулярный кислород. АТФ и НАДФ•H используются в реакциях темновой фазы фотосинтеза. |
49)Формы биологических связей в природе.Паразитизм как биологический феномен. Существуют две основные формы межвидовых взаимодействий: антибиоз и симбиоз. Антибиоз- невозможность существования двух видов организмов, основанная на конкуренции, прежде всего за источники питания (сапрофитные бактерии и плесневые грибы). Симбиоз- сожительство (греческое): - мутуализм - отношения между организмами разных видов полезны настолько, что раздельное существование невозможно (человек и микрофлора кишечника). Дисбактериоз – гибель нормальных бактерий и усиленное размножение бактерий, нечувствительных к антибиотику, и микроскопических грибов, которые могут явиться причиной заболевания; - комменсализм – форма симбиоза, при которой один вид использует остатки или излишки пищи другого, не причиняя ему видимого вреда (непатогенная ротовая и кишечная амеба в пищеварительной системе человека); - хищничество – пищевые взаимодействия при отсутствии пространственных взаимодействий; - паразитизм – форма межвидовых взаимоотношений, при которой один вид использует другой как источник питания и среду обитания (истинный, ложный, облигатный, облигатный, факультативный). По времени контакта : временные, постоянные (стационарные, периодические). По их локализации в организме хозяина: эктопаразиты (кровососущие насекомые и клещи), эндопаразиты (паразиты, обитающие в полостных органах, связанных с внешней средой (аскарида), и паразиты тканей внутренней среды (малярийный плазмодий)). |
27)Регуляция реакций фотосинтеза Эндогенные механизмы регуляции фотосинтеза. Реализация фотосинтетической функции растения в целом определяется с одной стороны значительной автономностью хлоропластов, а с другой – сложной системой связей фотосинтеза со всеми функциями растения. В ходе онтогенеза в растительном организме всегда присутствуют аттрагирующие зоны (зоны, притягивающие питательные вещества). В аттрагирующих центрах происходит либо новообразование и рост структур, либо интенсивный однонаправленный синтез запасных веществ (клубни, плоды и др.). В обоих случаях состояние аттрагирующих центров определяет величину «запроса» на фотосинтез. Если внешние условия не лимитируют фотосинтез, то ведущая роль принадлежит аттрагирующим центрам. Чем мощнее центры, аттрагирующие ассимиляты, тем интенсивнее фотосинтез. Второй. важный механизм регуляции фотосинтеза связан с фитогормонами и эндогенными ингибиторами роста и метаболизма. Фитогормоны образуются в разных частях растении, в том числе и хлоропластах, и действуют на процессы фотосинтеза как дистанционно, так и непосредственно на уровне хлоропластов. Дистанционное воздействие осуществляется благодаря регулирующему влиянию фитогормонов на процессы роста и развития, на отложение веществ в запас, на транспорт ассимилятов и т.д., т.е. на формирование и активность аттрагирующих центров. С другой стороны фитогормоны оказывают прямое действие на функциональную активность хлоропластов через изменение состояния мембран, активность ферментов, генерацию трансмембранного потенциала. Доказана также роль фитогормонов, в частности цитокинина, в биогенезе хлоропластов, синтезе хлорофиллов, ферментов ц.Кальвина.
На интенсивность фотосинтеза влияют такие факторы внешней среды, как: интенсивность и качество света, концентрация углекислого газа, температура, водный режим тканей растения, минеральное питание и др.
|
||
|
52)Борьба с малярией,задачи противомалярийной службы на современном этапе Лабораторный диагноз малярии можно поставить только в период, соответствующий стадии эритроцитарной шизогонии, когда в крови удается обнаружить паразитов. Профилактика малярии — раннее выявление и лечение больных, профилактическое лечение в зонах широкого распространения малярии. Как и при любых трансмиссивных заболеваниях, необходима прицельная борьба с переносчиками (осушение болот, опрыскивание химическими веществами место размножения переносчиков, средства индивидуальной защиты). |
56)Свиной цепень. Систематическое положение , морфология. Тип Plathelmintes класс Cestoidea Свиной цепень Taenia solium (рис. 20.8, В) — возбудитель тениоза и цистицеркоза. Этот паразит меньше предыдущего, он достигает в длину 3 м. На головке кроме присосок у него находится венчик из 22—32 крючьев. В гермафродитных члениках не две, а три дольки яичника; матка в зрелых члениках имеет не более 12 пар боковых ответвлений. Яйца не отличаются от яиц предыдущего вида. |
67)Эпистаз Эпистаз - взаимодействие генов, при котором активность одного гена находится под влиянием вариаций других генов. Ген, подавляющий фенотипические проявления другого, называется эпистатичным; ген, чья активность изменена или подавлена, называется гипостатичным. Примеры эпистатического влияния тесно связанных генов на приспособленность можно обнаружить в супергенах и главном комплексе гистосовместимости. Эффект может проявляться как напрямую на уровне генов, при этом продукт эпистатичного гена предотвращает транскрипцию гипостатичного, так и на уровне фенотипов. |
||
|
28)Хемосинтез(общие положения) способ автотрофного питания, при котором источником энергии для синтеза органических веществ служат процессы окисления различных неорганических веществ: аммиака, серы, водорода и соединений железа.Источником водорода д ля восстановления углекислого газа в ходе хемосинтеза являетсяя вода. Хемосинтез был открыт русским ученым Виноградским в 1887 .
|
30)Продуктивность гликолиза(по образованию молекул АТФ) 2 этап –гликолиз(бескислородный): осуществляется в гиалоплазме, с мембранами не связан,в нем участвуют ферменты,расщеплению подвергается глюкоза: --->60% теплота C6H12O6--->2C3H4O3+2H+E (пировиноград.ки-та) --->40% на синтез 2АДФ+2Ф=2АТФ+2НАД*Н
|
54)Кошачий сосальщик. Систематическое положение, морфология Класс Trematoda Кошачий сосальщик Opisthorchis felineus — возбудитель описторхоза. Длина тела до 13 мм. Характерная особенность—два хорошо окрашивающихся лопастевидных семенника на заднем конце тела. Яйца длиной 26—30 мкм, с крышечкой.
|
||
|
73)Хромосомные мутации ХРОМОСОМНЫЕ МУТАЦИИ - это перестройки хромосом, изменение их строения. Отдельные участки хромосом могут теряться, удваиваться, менять свое положение. Как и геномные мутации, хромосомные мутации играют огромную роль в эволюционных процессах.
Мутационная теория В 1898 г. русский ботаник Сергей Иванович Коржинский (1861 - 1900 гг.), а спустя два года нидерландский ботаник Хуго де Фриз (1848 - 1935 гг.) делают независимо друг от друга чрезвычайно важное генетическое обобщение, получившее название мутационной теории. Мутационная теория утверждает, что из двух категорий изменчивости - непрерывной и прерывистой, только последняя передаётся по наследству. Основные положения этой теории:
Основной ошибкой в мутационной теории Де Фриза было утверждение, что в результате мутации без участия естественного отбора могут сразу возникать новые виды. В действительности мутационная изменчивость наряду с комбинативной создаёт материал для естественного отбора, который формирует виды в процессе эволюции. В дальнейшем выяснилось, что ошибка Де Фриза была связана с тем, что растение энотера, с которым он работал, представляло собой сложную полигетерозиготу, а изменения, которые Де Фриз принял за мутации, - результат расщепления этой гетерозиготы. Тем не менее реальность мутационной изменчивости была в дальнейшем доказана многочисленными исследователями, и основные положения мутационной теории получили развитие и экспериментальные подтверждения.
|
74) Наследственные болезни человека, определяемые хромосомной трисомией Синдром Дауна - синдром трисомии 21 (1:700-1000). Описал Лэнгдон Даун . Для больных характерны округлой формы голова с уплощенным затылком, узкий лоб, широкое, плоское лицо. Типичны эпикант, запавшая спинка носа, косой (монголоидный) разрез глазных щелей, пятна Брушфильда (светлые пятна на радужке), толстые губы, утолщенный язык с глубокими бороздами, выступающий изо рта, маленькие, округлой формы, низко расположенные ушные раковины со свисающим завитком, недоразвитая верхняя челюсть, высокое нёбо, неправильный рост зубов, короткая шея. Синдром Патау - синдром трисомии 13 - встречается с частотой 1:6000. При СП наблюдаются тяжелые врожденные пороки. Дети с синдромом Патау рождаются с массой тела ниже нормы . У них выявляются умеренная микроцефалия, нарушение развития различных отделов ЦНС, низкий скошенный лоб, суженные глазные щели, расстояние между которыми уменьшено, микрофтальмия и колобома, помутнение роговицы, запавшая переносица, широкое основание носа, деформированные ушные раковины, расщелина верхней губы и нёба, полидактилия, флексорное положение кистей, короткая шея. У 80% новорожденных встречаются пороки развития сердца:Большинство больных с синдромом Патау (98%) умирают в возрасте до года. Синдром Эдвардса (СЭ)- синдром трисомии 18 - встречается с частотой примерно 1:7000. У девочек встречается значительно чаще, чем у мальчиков, что связано, возможно, с большей жизнестойкостью женского организма. Дети с трисомией 18 рождаются с низким весом, хотя сроки беременности нормальные или даже превышают норму. Фенотипические проявления синдрома Эдвардса многообразны. Наиболее часто отмечаются аномалии мозгового и лицевого черепа, мозговой череп долихоцефалической формы. Нижняя челюсть и ротовое отверстие маленькие. Глазные щели узкие и короткие. Ушные раковины деформированы и в подавляющем большинстве случаев расположены низко, несколько вытянуты в горизонтальной плоскости. Мочка, а часто и козелок отсутствуют. Наружный слуховой проход сужен, иногда отсутствует. Грудина короткая, из-за чего межреберные промежутки уменьшены и грудная клетка шире и короче нормальной. В 80% случаев наблюдается аномальное развитие стопы: пятка резко выступает, свод провисает (стопа-качалка), большой палец утолщен и укорочен. Продолжительность жизни детей с синдромом Эдвардса невелика: 60% детей умирают в возрасте до 3 мес, до года доживает лишь один ребенок из десяти; оставшиеся в живых - глубокие олигофрены.
|
40)Онтогенез и его периодизация. Прямое и непрямое развитие. онтогенез - индивидуальное развитие особи - начинается с момента слияния сперматозоида с яйцеклеткой и образования зиготы, заканчивается смертью. Есть два типа онтогенеза: прямой и непрямой. Непрямое развитие может быть личиночным, прямое развитие - в двух формах: неличиночной и внутриутробной. Личиночная форма характеризуется наличием одной или нескольких личиночных стадий. Личинки активно питаются, имеют временные органы. Этот тип развития сопровождается метаморфозом. Неличиночная форма развития характерна для рыб, рептилий, птиц. Яйца этих животных богаты желтком. Для дыхания, выделения, питания развивающегося зародыша есть временные органы. Внутриутробная форма характерна для млекопитающих и человека. Все функции зародыша осуществляются за счет организма матери, с помощью специального органа - плаценты. При любой форме онтогенеза выделяют следующие периоды: эмбриональный и постэмбриональный. До эмбрионального периода происходит гаметогенез - образование сперматозоидов и яйцеклеток. Эмбриональный период имеет следующие этапы: 1)Дробление - образование бластулы; 2)Гаструляция - образование зародышевых листков; 3)Гисто- и органогенез - образование органов и тканей зародыша. При личиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из яйцевых оболочек. При неличиночной форме онтогенеза эмбриональный период начинается с образования зиготы и заканчивается выходом из зародышевых оболочек. При внутриутробной форме онтогенеза эмбриональный период начинается с образования зиготы и длится до рождения.
|
||
|
87)Производные эктодермы Эпидермис кожи,ногти,волосы,потовые железы, вся нервная система: головной мозг,спинной мозг, ганглии, нервы;рецепторные клетки органов чувств, хрусталик глаза, зубная эмаль
|
88)Производные энтодермы Эпителий желудка, пищевода,кишечника, эпителий трахеи, бронхов,легких,печень,поджелудочная железа,эпителий желчного пузыря, щитовидная и паращитовидная железы,эпителий мочевого пузыря,эпителий мочеиспускательного канала,хорда
|
89)Производные мезодермы Гладкая мускулатура всех органов,скелетная мускулатура,сердечная мышца, соединительная ткань,кости,хрящи,дентин зубов,кровь,кровеносные сосуды,брыжейка,почки,семенники и яичники
|
||
|
83)Мутационный процесс и его значение для видообразования Понятие о мутациях было введено в науку голландцем де Фризом. Мутации - это наследственные изменения, приводящие к увеличению или уменьшению количества генетического материала, к изменению нуклеотидов или их последовательности. Модификационная изменчивость отражает изменения фенотипа, не затрагивая генотипа. Противоположной ей является другая форма изменчивости - генотипическая, или мутационная (по Дарвину - наследственная, неопределенная, индивидуальная), меняющая генотип. Мутация - стойкое наследственное изменение генетического материала. Отдельные изменения генотипа называются мутациями. В понятие наследственной изменчивости входят генотипическая и цитоплазматическая изменчивость. Первая делится на мутационную, комбинативную, соотносительную. Комбинативная изменчивость возникает при кроссинговере, независимом расхождении хромосом в мейозе и случайном слиянии гамет при половом размножении. В состав мутационной изменчивости входят геномные, хромосомные и генные мутации. Термин мутация был введен в науку Г. де Фризом. Его биография и основные научные достижения располагаются в разделе. Геномные мутации связаны с возникновением полиплоидов и анэуплоидов. Хромосомные мутации определяются межхромосомными изменениями - транслокацией или внутрихромосомными перестройками: делецией, дупликацией, инверсией. Генные мутации объясняются изменениями в последовательности нуклеотидов: увеличением или уменьшением их числа (делеция, дупликация), вставкой нового нуклеотида или поворотом участка внутри гена (инверсия). Цитоплазматическая изменчивость связана с ДНК, которая находится в пластидах и митохондриях клетки. Наследственная изменчивость родственных видов и родов подчиняется закону гомологических рядов Вавилова. Большинство возникающих мутаций рецессивны и неблагоприятны для организма, даже могут вызвать его гибель. В сочетании с аллельным доминантным геном рецессивные мутации не проявляются фенотипически. Мутации имеют место в половых и в соматических клетках. Если мутации происходят в половых клетках, то они называются генеративными и проявляются в том поколении, которое развивается из половых клеток. Изменения в вегетативных клетках называются соматическими мутациями. Такие мутации приводят к изменению признака только части организма, развивающегося из измененных клеток. У животных соматические мутации не передаются последующим поколениям, поскольку из соматических клеток новый организм не возникает. Иначе у растений: в гибридных клетках растительных организмов репликация и митоз могут осуществляться в разных ядрах несколько по-разному. На протяжении ряда клеточных генераций происходит потеря отдельных хромосом и отбираются определенные кариотипы, способные сохраняться в течение многих поколений. |
86)Понятие о ткани. Типы тканей Ткань — система клеток и межклеточного вещества, объединённых общим происхождением, строением и выполняемыми функциями. Строение тканей живых организмов изучает наука гистология. Совокупность различных и взаимодействующих тканей образуют органы. Соединительные виды тканей и их функции Все ткани этой группы характеризуются большим количеством межклеточного вещества. В некоторых случаях оно выступает в жидком агрегатном состоянии, в некоторых — в жидком, иногда — в виде аморфной массы. К этой группе принадлежат семь типов. Это плотная и рыхлая волокнистые, костная, хрящевая, ретикулярная, жировая, кровь. В первой разновидности преобладают волокна. Она расположена вокруг внутренних органов. Ее функции заключаются в придании им эластичности и их защите. В рыхлой волокнистой ткани аморфная масса преобладает над самими волокнами. Она полностью заполняет промежутки между внутренними органами, в то время как плотная волокнистая формирует только своеобразные оболочки вокруг последних. Она также играет защитную роль.
Мышечные ткани Данная группа подразделяется на три вида. Мышца бывает гладкая, поперечно-полосатая и сердечная. Все мышечные ткани похожи тем, что состоят из длинных клеток - волокон, в них содержится очень большое количество митохондрий, так как им необходимо много энергии для осуществления движений. Гладкая мышечная ткань выстилает полости внутренних органов. Сокращение таких мышц мы не можем контролировать сами, так как они иннервируются автономной нервной системой.
Эпителиальная Это такие виды тканей, клетки которых плотно прилегают друг к другу. Они могут иметь разнообразную форму, но всегда расположены близко. Все различные виды тканей данной группы имеют сходство и в том, что межклеточного вещества в них мало. Оно в основном представлено в виде жидкости, в некоторых случаях его может и не быть. Это виды тканей организма, которые обеспечивают его защиту, а также выполняют секреторную функцию.
Нервная ткань Так как она бывает только одной разновидности, начнем с нее. Клетки данной ткани называются нейронами. Каждый из них состоит из тела, аксона и дендритов. Последние - это отростки, по которым электрический импульс передается от клетки к клетке. Аксон у нейрона один - это длинный отросток, дендритов несколько, они более мелкие, чем первый. В теле клетки находится ядро. Кроме того, в цитоплазме расположены так называемые тельца Ниссля - аналог эндоплазматического ретикуллума, митохондрии, которые вырабатывают энергию, а также нейротрубочки, которые участвуют в проведении импульса от одной клетки к другой. |
48)Регенрация и ее типы Во всех периодах онтогенеза организмы способны к восстановлению утраченных или поврежденных частей тела. Процессы, направленные на восстановление разрушенных биологических структур, называются регенерацией. Регенерация может быть физиологической и репаративной. Физиологическая регенерация свойственна всем организмам. Она связана с восстановлением утраченных структур в процессе жизнедеятельности организма. Регенерация может осуществляться на клеточном, тканевом, органном уровнях. Например, у членистоногих физиологическая регенерация связана с линькой. У млекопитающих и человека систематически слущивается кожный эпителий, быстро происходит смена эритроцитов и др. тканей. Репаративная регенерация - это восстановление части тела организма, отторгнутой насильственным путем. Способность к регенерации выражена по-разному. Одни животные способны восстанавливать целостный организм из отдельных клеток. Хорошо выражена репаративная регенерация у кишечнополостных (гидра), ресничных червей (планария). Другие животные восстанавливают только нарушенные органы. Ракообразные способны восстанавливать утраченные конечности, антенны. Хвостатые амфибии и личинки бесхвостых восстанавливают конечности, хвост. Некоторые животные могут восстанавливать внутренние органы. При этом восстанавливается не форма, а масса органа. Сначала заживляется рана, а затем происходит увеличение оставшейся части органа за счет размножения клеток. Например, так регенерирует печень. Иногда регенерация сводится лишь к рубцеванию ран. У человека может регенерировать эпителиальная, мышечная, соединительная, кожные ткани, периферические нервы. Чаще всего регенерация у..Млекопитающих приводит к заживлению ран, что препятствует проникновению болезнетворных микробов в организм. При повышении физиологической нагрузки иногда происходит увеличение органов и тканей. Возникает гипертрофия органов, например, гипертрофия желудочков сердца, или при удалении одной почки, вторая почка может увеличиться в размерах. Процессы, происходящие при регенерации, регулируются нервной системой и гуморально. Знание процессов регенерации необходимо в хирургической практике.
|
||
|
41)Эмбриогенез:дробление В результате оплодотворения образуется зигота, которая начинает дробиться. Дробление сопровождается митотическим делением. Нет роста клеток, и объем зародыша не изменяется. Это происходит потому, что между делениями в короткой интерфазе отсутствует постмитотический период, а синтез ДНК начинается в телофазе предшествующего митотического деления. Клетки, образующиеся в процессе дробления, называются бластомерами, а зародыш -бластулой. Типы дробления зависят от количества и распределения желтка в яйцеклетках Дробление может быть: •полным равномерным; •полным неравномерным; •неполным дискоидальным; •неполным поверхностным. Полное равномерное дробление характерно для изолецитальных яиц, например, для ланцетника. Ядро зиготы делится митозом на два, затем делится цитоплазма. Борозда дробления проходит по меридиану, образуя два бластомера. Затем снова делится ядро, и на поверхности зародыша появляется вторая борозда дробления, идущая по меридиану перпендикулярно первой. Образуются четыре бластомера. Третья борозда проходит по экватору и делит его на восемь частей. Затем происходит чередование меридионального и экваториального дроблений. Число бластомеров увеличивается. Зародыш на стадии 32 бластомеров называют морулой. Дробление продолжается до образования зародыша, похожего на пузырек, стенки которого образованы одним слоем клеток, называемом бластодермой. Бластомеры расходятся от центра зародыша, образуя полость, которая называется первичной или бластоцелью. Бластомеры имеют одинаковые размеры. В результате такого дробления образуется целобластула Полное неравномерное дробление характерно для телолецитальных яиц с умеренным содержанием желтка, например, у лягушки. Первая и вторая борозды дробления проходят по меридианам и полностью делят яйцо на четыре части. Третья борозда смещена в сторону анимального полюса, где нет желтка. Бластомеры имеют неодинаковую величину: на анимальном полюсе они меньше (микромеры), на вегетативном больше (макромеры). Желток затрудняет дробление, и поэтому дробление макромеров идет медленнее, чем микромеров. Стенка бластулы состоит из нескольких рядов клеток. Первичная полость мала и смещена к анимальному полюсу. Образуется амфибластула. Неполное дискоидальное дробление характерно для телолецитальных яиц с большим содержанием желтка, например, для рептилий, птиц. Дробление идет только на анимальном полюсе. Первая и вторая борозды дробления проходят по меридиану перпендикулярно друг другу. Третья борозда смещена к анимальному полюсу, в результате чего образуется зародышевый диск. Бластоцель располагается под слоем бластодермы в виде щели. Бластула называется дискобластулой Неполное поверхностное дробление характерно для центролецитальных яиц, например, для членистоногих. Ядра центролецитальных яиц многократно делятся и перемещаются к периферии, где в цитоплазме нет желтка. Образуются бластомеры. Бластула имеет один слой бластомеров. Бластоцель заполнена желтком. Такая бластула называется перибластулой.
|
46)Эмбриональное развитие и апоптоз Эмбриональное развитие - развитие организма животного, происходящее в оболочках яйца вне материнского организма или внутри него. Эмбриональное развитие предшествует период предзародышевого развития, когда растет, формируется и созревает яйцо. В ходе эмбрионнального развития из одной относительно просто организованной яйцеклетки образуется многоклеточный организм, состоящий из различных органов и тканей и способный к самостоятельному существованию. У некоторых животных, например иглокожих, зародыши выходят из оболочек на очень ранних стадиях, и основные процессы развития проходят у них в постэмбриональный период. У всех животных эмбриональное развитие складывается из оплодотворения или (при партеногенезе) активации яйца, делений дробления, гаструляции, органогенеза и выхода из оболочек или рождения. Апоптоз – генетически запрограммированная гибель клеток, благодаря которой в ходе эмбриогенеза происходит исчезновение ненужных органов или частей органов. В целом эмбриональное развитие происходит с образованием большого избытка клеток, которые своевременно подвергаются апоптозу. Это создает запас клеточного материала и возможность последовательного развития. В эмбриогенезе одновременно с ростом тканей происходит регрессия частей эмбриональных зачатков, тканей и органов, инволюция провизорных (временных) органов, появление просвета в полых органах. Механизм апоптоза заключается в том, что в отличие от некроза – патологической массивной гибели клеток, расположенных в одном участке ткани, при апоптозе гибнут единичные клетки, расположенные в ткани мозаично, их гибель происходит запрограммированно в результате воздействия специально синтезируемых белков апоптоза. Под действием белков апоптоза происходит упорядоченное расщепление ДНК, в отличие от некроза, где расщепление ДНК незакономерное, случайное. Таким образом, апоптоз представляет собой регуляцию числа клеток в развивающихся тканях, участвует в перестройке тканей, в исчезновении рудиментальных зачатков, в элиминации дефектных клеток. Наиболее выражены процессы апоптоза в нервной системе, где в ходе гистогенеза гибнет до 85 % всех нейронов. Все морфологические процессы находятся под контролем семейства генов, которые называются гомеозисными генами. В геноме млекопитающих их 38. Они сгруппированы в четыре комплекса и расположены в определенных хромосомах. Гомеозисные гены содержат гомеобокс (участок ДНК, содержащий определенное количество пар нуклеотидов), который обусловливает одинаковую функцию генов, поэтому гомеозисные гены обеспечивают развитие признаков, общих для всех видов животных (образование зародышевых листков, осей, нервной системы и т. д.).
|
59)Аскарида,систематическое положение,морфология. К нематодам относится аскарида человеческая (Ascaris lumbricoides), паразитирующая в тонком кишечнике человека и вызывающая заболевание -аскаридоз. Аскаридами заражено около 1 млрд. человек на всем земном шаре. Аскарида человеческая - крупный гельминт красновато-желтого цвета. Имеет вытянутое тело веретенообразной формы с заостренными концами. Самки крупнее самцов. Длина самки до 40 см, самцов - до 25 - 30 см. У самок задний конец прямой, у самцов -заострен и загнут на брюшную сторону. Тело аскариды покрыто кожно-мускульным мешком, образованным кутикулой, гиподермой и одним слоем мышц. Кутикула - плотная оболочка, выполняющая функции наружного скелета и защищающая паразита от внешних воздействий. Внутри кожно-мускульного мешка имеется первичная полость, заполненная ядовитой жидкостью, представляющая жидкие продукты жизнедеятельности аскариды. Жидкость находится под большим давлением и образует гидроскелет. В полости тела расположены внутренние органы . На переднем конце тела аскариды имеется ротовое отверстие, окруженное тремя губами. Ротовое отверстие ведет в мускулистую глотку, а затем в пищевод. Глотка и пищевод эктодермального происхождения. За пищеводом следует кишечник энтодермального происхождения. Кишечник заканчивается анальным отверстием. Дыхательная и кровеносная системы у аскариды отсутствуют. Выделительная система состоит из одной кожной железы, находящейся в передней части тела аскариды. Нервная система состоит из окологлоточного нервного кольца и отходящих от него нервных стволов. Органы чувств у аскариды развиты слабо. Половая система имеет трубчатое строение. У самки половая система состоит из парных яичников, парных яйцеводов, представляющих собой трубки большого диаметра, и парных маток, имеющих еще больший диаметр и соединяющихся в непарное влагалище, открывающееся отверстием в верхней трети тела аскариды на брюшной стороне. Половая система самца непарная, имеет тонкий трубчатый семенник, семяпровод и семяизвергательный канал, открывающийся в заднюю кишку.
|
||
|
61)Острица. Систематическое положение, морфология:цикл развития. Пути заражения хозяев.Лабораторная диагностика,профилактика заболевания. К круглым червям относят острицу (Enterobins vermicularis). Паразитирование остриц у человека так же широко распространено, как аскарид. Заболевание, вызываемое паразитированием остриц, называют энтеробиозом. Взрослые черви имеют небольшие размеры: самки – 9 - 12 мм, самцы – 3 - 5 мм. Передний конец остриц имеет небольшие кутикулярные крылья головной везикулы. Задний конец тела самца крючковидно изогнут и снабжен двумя боковыми крыловидными пластинками. Пищевод имеет расширение - бульбус. Паразитирует острица в кишечнике человека. Яйца, откладываемые самкой, созревают во внешней среде в течение 4-6 часов. С грязными руками яйца попадают через рот и в тонком кишечнике происходит выход личинок из яйцевых оболочек. Личинки мигрируют в начальные отделы толстой кишки. Через 12-14 дней они достигают половой зрелости. Взрослые особи живут в кишечнике около 30 дней. Для откладывания яиц самки ночью выползают из анального отверстия и вызывают сильный зуд. При расчесывании зудящих мест яйца остаются под ногтями. С этим связано широкое распространение энтеробиоза, особенно среди детей . Для постановки диагноза необходимо сделать соскоб с перианальных складок кожи. Чтобы избавиться от энтеробиоза, надо соблюдать правила личной гигиены. Заболевание, вызываемое паразитированием власоглава (Trichocephaltis trichiurus), называют трихоцефалезом. Паразитирует власоглав в слепой кишке, начальном отделе толстой кишки у человека. Гельминт имеет тонкую волосовидную переднюю часть тела (откуда произошло название паразита) и более широкую заднюю. Длина самца 30 - 45 мм, самки - 35-55 мм. Заражение трихоцефалезом происходит, если человек не соблюдает правила личной гигиены. С немытыми овощами, фруктами и грязными руками инвазионные яйца попадают в рот. Из яиц выходят личинки, которые мигрируют по кишечнику и достигают слепой кишки. Передним концом прикрепляются к слизистой оболочке. Половой зрелости власоглавы достигают через месяц после заражения. Яйца, выделяемые больными трихоцефалезом, имеют вытянутую форму, на полюсах яиц располагаются прозрачные пробочки. Во внешней среде внутри яиц при 24 - 28°С через 4 недели формируются инвазионные личинки. Цикл развития происходит без смены хозяев. У человека власоглав может паразитировать около 5 лет .Диагноз ставят при обнаружении яиц власоглава в фекалиях. Профилактика связана с соблюдением правил личной гигиены и охраной окружающей среды от заражений нечистотами. |
76)Синдром «кошачьего крика» Синдром кошачьего крика (синонимы: болезнь кошачьего крика, синдром Лежена по имени описавшего его в 1963 г. французского ученого) . Генетика Кариотип 46 XX или ХУ, 5р-. Диагноз подтверждается кариологическим исследованием с применением одного из методов идентификации хромосом. Хромосомно синдром кошачьего крика объясняется частичной моносомией; он развивается при делеции (с утратой от трети до половины, реже полная утрата) короткого плеча пятой хромосомы. Для развития клинической картины синдрома имеет значение не величина утраченного участка, а конкретный незначительный фрагмент хромосомы. Изредка отмечается мозаицизм по делеции или образование кольцевой хромосомы-5. Клиника При этом синдроме наблюдается: общее отставание в развитии, низкая масса при рождении и мышечная гипотония, лунообразное лицо с широко расставленными глазами характерный плач ребёнка, напоминающий кошачье мяуканье, причиной которого является изменение гортани (сужение, мягкость хрящей, уменьшение надгортанника, необычная складчатость слизистой оболочки) или недоразвитие гортани. Признак исчезает к концу первого года жизни. Кроме того, встречаются врожденные пороки сердца, костно-мышечной системы и внутренних органов, микроцефалия, птоз, низкое расположение и деформация ушных раковин, кожные складки впереди уха, гипертелоризм (увеличенное расстояние между какими-либо парными органами или анатомическим образованиями (например, между внутренними краями глазниц, грудными сосками) , эпикантус (поперечная кожная складка около внутреннего угла глаза, обычно двусторонняя; наиболее чётко выражена при болезни Дауна, антимонголоидный разрез глаз. Частота синдрома примерно 1:45000. Соотношение полов М1 : Ж1,3. Клиническая картина синдрома и продолжительность жизни людей с этим синдромом довольно сильно варьирует по сочетанию врождённых пороков развития органов. Лечение симптоматическое. Показаны средства, стимулирующие психомоторное развитие, лечебный массаж и гимнастика.
|
36)Половое размножение, его способы и биологическое значение. Половое размножение –— воспроизведение себе подобных, происходящее, как правило, с участием двух особей в результате слияния гамет, т. е. копуляции яйцеклетки и сперматозоида. Яйцеклетки образуются у особей женского пола (материнский организм), сперматозоиды — у особей мужского пола (отцовский организм). Половое размножение свойственно как растительным, так и животным организмам. У растений яйцеклетки образуются в специальных органах — архегониях, сперматозоиды — в антеридиях. У животных яйцеклетки формируются в яичниках, сперматозоиды — в семенниках. Разница заключается в том, что у животных образованию половых клеток (гамет) предшествует мейоз, а у растений мейоз происходит перед образованием спор, из которых развиваются заростки. На них формируются архегонии с яйцеклетками и антеридии со сперматозоидами. Таким образом, у любых организмов — растений или животных — гаметы обязательно гаплоидные, а зигота диплоидная, из нее формируется диплоидный зародыш, половина хромосом которого от материнского организма, а половина от отцовского. Объединение генетической информации может происходить при конъюгации (временном соединении особей для обмена информацией, как это происходит у инфузорий) и копуляции (слиянии особей для оплодотворения) у одноклеточных животных, а также при оплодотворении у представителей разных царств. Особым случаем полового размножения является партеногенез у некоторых животных (тли, трутни пчел). В этом случае новый организм развивается из неоплодотворенного яйца, но до этого всегда происходит образование гамет. Половое размножение у покрытосеменных растений происходит путем двойного оплодотворения. Дело в том, что в пыльнике цветка образуются гаплоидные пыльцевые зерна. Ядра этих зерен делятся на два – генеративное и вегетативное. Попав на рыльце пестика, пыльцевое зерно прорастает, образуя пыльцевую трубку. Генеративное ядро делится еще раз, образуя два спермия. Один из них, проникая в завязь, оплодотворяет яйцеклетку, а другой сливается с двумя полярными ядрами двух центральных клеток зародыша, образуя триплоидный эндосперм. Все вышеперечисленное справедливо только для эукариот. У прокариот тоже есть половое размножение, но происходит оно по-другому. Таким образом, при половом размножении происходит смешивание геномов двух разных особей одного вида. Потомство несет новые генетические комбинации, что отличает их от родителей и друг от друга. Различные комбинации генов, проявляющиеся в потомстве в виде новых, интересующих человека признаках, отбираются селекционерами для выведения новых пород животных или сортов растений. В некоторых случаях применяют искусственное оплодотворение. Это делается и для того, чтобы получить потомство с заданными свойствами, и для того, чтобы преодолеть бездетность некоторых женщин. |
||
|
44)Эмбриогенез: эмбриональная индукция Эмбриональная индукция — это взаимодействие частей развивающегося зародыша, при котором один участок зародыша влияет на судьбу другого участка. Явление эмбриональной индукции с начала XX в. изучает экспериментальная эмбриология.
|
34)Биологический смысл мейоза биологический смысл мейоза заключается в сохранении постоянства хромосомного набора для данного вида. Кроме того, при мейозе, в отличие от митоза, происходит частичная перекомбинация наследственной информации, возникают новые сочетания генов. Это повышает выживаемость вида в процессе эволюции. Мейоз включает два деления клетки, которые также состоят из профазы, метафазы, анафазы и телофазы. Перед началом мейоза, как и при митозе, ДНК клетки удваивается. Каждая хромосома теперь состоит из двух дочерних хроматид. В профазе мейоза (как и при митозе) хромосомы становятся видимыми, центриоли расходятся к полюсам, образуется веретено деления.
|
47)Эмбриогенез человека Эмбриогенез человека - это часть его индивидуального развития, онтогенеза. Он тесно связан с прогенезом (образованием половых клеток и ранним постэмбриональным развитием. Эмбриология человека изучает процесс развития человека, начиная с оплодотворения и до рождения. Эмбриогенез человека, продолжающийся в среднем 280 суток (10 лунных месяцев ), подразделяется на три периода: начальный (первая неделя развития), зародышевый (вторая-восьмая недели), и плодный (с девятой недели до рождения ребенка).
|
||
|
80)Цитологический метод диагностики хромосомных нарушений у человека основан на микроскопическом изучении хромосом в клетках человека. Современный этап в применении цитогенетического метода связан с методом дифференциального окрашивания хромосом. Применение цитогенетического метода позволяет изучать нормальную морфологию хромосом и кариотипа в целом, определять генетический пол организма, диагностировать различные хромосомные болезни, связанные с изменением числа хромосом или с нарушением их структуры, изучать процессы мутагенеза на уровне хромосом и кариотипа. Применение: пренатальная диагностика хромосомных болезней, что дает возможность путем своевременного прерывания беременности предупредить появление потомства с грубыми нарушениями развития. Материал: лимфоциты периферической крови, клетки костного мозга, фибробласты, клетки опухолей и эмбриональных тканей и др. Непременным требованием для изучения хромосом является наличие делящихся клеток. В качестве экспресс-метода, выявляющего изменение числа половых хромосом, используют метод определения полового хроматина в неделящихся клетках слизистой оболочки щеки. Половой хроматин, или тельце Барра, образуется в клетках женского организма одной из двух Х-хромосом. Оно выглядит как интенсивно окрашенная глыбка, расположенная у ядерной оболочки. При увеличении количества Х-хромосом в кариотипе организма в его клетках образуются тельца Барра в количестве на единицу меньше числа хромосом. При уменьшении числа Х-хромосом (моносомия X) тельце Барра отсутствет. В мужском кариотипе У-хромосома может быть обнаружена по более интенсивной по сравнению с другими хромосомами люминесценции при обработке их акрихинипритом и изучении в ультрафиолетовом свете. |
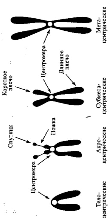
81)Типы хромосом 1)Метацентрические - центромера располагается строго по середине. 2)Субметацентрические - центромера смещена. 3)Акроцентрические - центромера сильно смещена. 4)Телоцентрические - нет 1 пары плеч.
|
84)Естественный отбор, его формы и значения для видообразования Популяции человека — сообщества, внутри которых браки заключаются чаще, чем с людьми других популяция различия между популяция всегда имеют групповой характер, касаясь частоты и географического распространения некоторых морфологических, физиологических и генетических признаков, в том числе вредных мутаций, что очень важно для генетики медицинской и изучения наследственных заболеваний. В человеческом обществе популяция включены в систему качественно своеобразных, специфических для людей социальных структур. В связи с этим общее направление, темпы и конкретные формы истории популяция находятся в зависимости от закономерностей социально-экономического развития, оказывающего мощное воздействие на все факторы микроэволюции: частоту мутаций, периодические колебания численности индивидов в популяция, характер изоляции и обусловленных ею границ между популяция и, наконец, на естественный отбор, роль которого непрерывно снижалась в процессе антропогенеза. На основе сложного взаимодействия смежных популяция складывались человеческие расы. На смену географических изоляции, игравшей главную роль на ранних этапах истории популяция человека, приходит изоляция общественная, этно-языковая, классово-сословная, производственно-профессиональная, государственно-политическая, религиозно-конфессиональная и др. В силу этого границы, разделяющие популяция у людей, часто совпадают с границами между теми или иными социальными общностями, в первую очередь между народами, этносами.
|
||
|
33)Амитоз Амитоз — прямое (простое) деление интерфазного ядра путем перетяжки. Происходит вне митотического цикла, т. е. не сопровождается сложной перестройкой всей клетки; спирализации хромосом также не происходит. Понятно, что при этом не обеспечивается равномерное распределение генетического материала между дочерними ядрами. Амитоз может сопровождаться делением клетки, а может ограничиваться лишь делением ядра без разделения цитоплазмы, что приводит к образованию дву- и многоядерных клеток. Клетка, претерпевшая амитоз, в дальнейшем неспособна вступить в нормальный митотический цикл. По сравнению с митозом амитоз встречается довольно редко. В норме он наблюдается в высокоспециализированных тканях, в клетках, которым уже не предстоит делиться: в эпителии и печени позвоночных, в зародышевых оболочках млекопитающих, в клетках эндосперма семени растений. Амитоз наблюдается также при необходимости быстрого восстановления тканей (после операций и травм). Амитозом также часто делятся клетки злокачественных опухолей.
|
90)Теория канцерогенеза. Канцерогенные факторы Канцерогенез – комплексный многоступенчатый процесс, включающий изменения не менее чем в 10 генетических факторах, каждый из которых является скоростьлимитарующим. В организме носителя каждая стадия процесса представляет собой физиологический барьер, который должен быть преодолен клеткой, прогрессирующей в сторону малигнизации (злокачественная трансформация). Существование множественности барьеров указывает на то, что малигнизация – явление редкое. В организме человека 1015 клеток. В течение жизни происходит их обновление в объеме, равном 10 объемам человеческого тела. Из этого становится понятным, что только тонкая сбалансированность процессов пролиферации, дифференцировки и апоптоза позволяет поддерживать нормальное развитие и функционирование всех органов и тканей. Пролиферация обеспечивает воспроизведение клеток, дифференцировка – приобретение ими индивидуальных черт и способности к специализированным видам деятельности, а апоптоз – разрушение старых и поврежденных клеток. Рак представляет собой совокупность генных болезней, характеризующихся неконтролируемой клеточной пролиферацией. В настоящее время доказано, что нарушения, ответственные за развитие опухолей, происходят на уровне ДНК. За исключением вирусиндуцированных, которые у людей достаточно редки. Трансформация клеток в раковые является результатом структурных изменений в специфических генах, кодирующих белки, принимающие участие в регуляции роста, деления и гибели клеток.
|
45) Постэмбриональное развитие Постэмбриональное развитие (от лат. post — после и греч. émbryon — зародыш) , послезародышевое развитие, период онтогенеза многоклеточных животных, следующий за периодом зародышевого развития и заканчивающийся обычно наступлением половой зрелости и (у большинства животных) прекращением роста. П. р. начинается после выхода зародыша из яйцевых и зародышевых оболочек, когда организм становится способным к активному питанию и перемещению. При переходе к П. р. организм или с самого начала обладает основными морфологически признаками половозрелой особи (прямое развитие) , или существенно отличается от неё, и вылупляющаяся из яйца личинка переходит к взрослому состоянию посредством превращения, или метаморфоза. В период П. р. продолжается рост, происходит дальнейший органогенез, гистогенез, усложняются функции развивающегося организма; особенно характерно установление окончательных пропорций тела. У некоторых животных П. р. составляет большую часть жизни. Так, среди насекомых у ряда цикад личинка живёт 17 лет, а половозрелое насекомое — одно лето; личинка подёнки живёт до 3 лет, а половозрелая особь — часто 1 сутки. Необходимость защиты от врагов и активного добывания пищи обеспечивается в период П. р. рядом приспособлений к самостоятельному образу жизни, сохраняющихся иногда в течение всей жизни животного, а иногда — при непрямом, или личиночном, развитии — только в течение П. р. (т. н. провизорные органы).
|
||
|
50)Малярийные плазмодии,систематическое положение, видовые различия. Малярийный комар-переносит возбудителей малярийного плазмодия. Малярийные комары,их личинки и куколки отличаются от обыкновенных. Длина и форму щупиков различны и это отличительные особенности комаров. У самок обыкновенных комаров нижнечелюстные щупики в 3 раза короче, чем хоботок, а у малярийных - равны хоботку. У самцов комара обыкновенного нижнечелюстные щупики длиннее хоботка, не имеют утолщений на концах; у самцов малярийного комара щупики по длине равны хоботку и имеют утолщения на концах. Яйца, отложенные самкой малярийного комара на поверхности воды плавают по одиночке. Для этого они имеют воздушные камеры. Самки обыкновенного комара откладывают яйца на воду, склеивая их и образуя "лодочку". Из яиц выходят личинки. Личинки малярийного комара на спинной стороне предпоследнего членика имеют пару стигм. Эти личинки располагаются параллельно поверхности воды. Живут в чистых водоемах. Личинки обыкновенного комара на предпоследнем членике брюшка имеют дыхательный сифон в виде узкой трубочки со стигмой на свободном конце. Личинки обыкновенных комаров располагаются под углом к поверхности воды, прикрепляясь концом сифона. Личинки усиленно питаются и растут, 4 раза линяют и превращаются в стадию куколки. Куколки не питаются. На стадии куколки происходит перестройка внутренних органов и появляются органы взрослого комара. Куколки комара подвижны и могут плавать. Тело куколки состоит из широкой головогруди и узкого брюшка из 9 сегментов. У малярийного комара на спинной стороне головогруди расположена пара дыхательных трубочек, расширенных вверху в виде воронок, у обыкновенного комара дыхательные трубочки имеют цилиндрическую форму. Из куколок вылупляются комары, сначала бесцветные. Через несколько минут крылья комаров расправляются, приобретают нормальную окраску и насекомые слетают с поверхности воды. При посадке брюшко малярийного комара приподнято и находится под углом к поверхности, а у обыкновенного комара при посадке брюшко находится параллельно поверхности . Продолжительность жизни самки в теплое время года до 3 месяцев. Уничтожают комаров на всех стадиях развития. Для избавления от взрослых форм применяют различные инсектициды. Царство Животные П/царство Одноклеточные (Простейшие) Н/тип Беспозвоночные (Invertebrata) Тип Споровики (Sporozoa) Отряд Кокцидии (Coccidia) Подотряд Кровяные споровики Род Плазмодий (Plasmodium) Вид Малярийный плазмодий (Plasmodium malariae)
|
58)Круглые черви. Общая эколого-морфилогическая характеристика типа. Особеннсоти органиации.Важнейшие представители. Значение для медицины Тип Круглые черви (нематоды) насчитывают около 500 000 видов. Представители этого типа обитают в морях, пресных водоемах, в почве. Многие круглые черви приспособились к паразитическому образу жизни. Они могут паразитировать у беспозвоночных и позвоночных животных, у растений и у человека. Развитие круглых червей происходит из трех зародышевых листков. Стенки внутренней полости не выстланы эпителием. Имеется первичная полость тела и кожно-мускульный мешок. Характерна билатеральная симметрия. Тело нематод круглое (в поперечном сечении), удлиненное, веретеновидное, передний и задний концы заострены. Для круглых червей характерно наличие мышечной, выделительной, пищеварительной, нервной и половой систем органов. Нематоды раздельнополы (есть самцы и самки). Пищеварительная система заканчивается анальным отверстием. С медицинской точки зрения, следует выделить еще одну группу нематод, неспецифических паразитов человека, которые осуществляют лишь миграцию в его организме, не достигая половой зрелости. Болезни, вызываемые круглыми червями, называют нематодозами. Круглых червей, сохранивших связь с внешней средой, яйца или личинки которых развиваются в почве, называют геогельминтами. Более специализированных паразитов, развивающихся с участием промежуточных хозяев, называют биогельминтами. Геогельминты обитают в просвете кишки и размножаются яйцами, которые выводятся с фекалиями и развиваются далее в почве. Они либо сами через определенное время становятся инвазионными, либо из них развиваются личинки, ведущие некоторое время свободный образ жизни и позже становящиеся инвазионными. Геогельминты, поражающие человека, не могут паразитировать у животных. Соответственно этому нематодозы, вызываемые этими паразитами, являются антропонозными болезнями. Заражение большей частью геогельминтов осуществляется при проглатывании яиц или личинок с продуктами, загрязненными почвой. Для диагностики всех нематодозов этой группы важно обнаружение яиц в фекалиях больного. Профилактические меры направлены на предотвращение попадания инвазионных яиц в пищеварительную систему—личная гигиена и гигиена питания, реже — другие меры. Часть геогельминтов, попадая в пищеварительную систему человека, быстро достигают половой зрелости и начинают размножаться в кишечнике, не мигрируя по организму хозяина. Личинки других перед достижением половой зрелости обязательно передвигаются по кровеносным сосудам и дыхательной системе и только после этого развиваются в кишечнике. |
60)Цикл развития,пути заражения хозяина. Лабораторная диагностика и профилактика аскаридоза Оплодотворение яиц происходит в матке. Самка откладывает более 200 000 яиц в сутки. Яйцам аскариды для развития необходимо попасть во внешнюю среду, где есть кислород, определенная влажность и температура 24 - 30°C. Яйца покрыты пятью оболочками, защищающими развивающийся зародыш от неблагоприятных условий (рис. 384). При благоприятных условиях в течение 15 - 20 дней в яйце формируется личинка, способная к дальнейшему развитию в организме человека. Такое яйцо называют инвазионным. В почве яйца могут сохранять инвазионность до 10 лет. Заражение людей аскаридозом происходит при употреблении в пищу ягод, овощей, загрязненных инвазионными яйцами аскариды при удобрении огородов и ягодников необезвреженными фекалиями человека. Цикл развития аскариды идет без смены хозяев. Из проглоченных яиц в кишечнике человека выходят личинки. С током крови они проходят в печень, через полую вену в правое предсердие, правый желудочек и по легочной артерии в альвеолы легких, где находится в среде, богатой кислородом. Затем они поднимаются по дыхательным путям в глотку, попадают в рот, вторично проглатываются и оказываются в кишечнике. Миграция личинок продолжается 9 - 12 дней. За это время личинки растут, несколько раз линяют, у них изменяется обмен веществ. Попав вторично в кишечник, личинки в течение 2,5 - 3 месяцев растут и превращаются во взрослых аскарид, способных выделять яйца. Продолжительность жизни аскариды в кишечнике около года Болезнь, вызванная паразитированием аскарид у человека называется аскаридозом. Мигрирующие личинки травмируют стенки кишечника, печень, легкие. При сильной инвазии могут возникать пневмонии, иногда бывают признаки аллергии: крапивница, насморк, кашель. Паразитирование взрослых аскарид в кишечнике связано с общей интоксикацией организма, кишечными расстройствами. Чтобы не заразиться аскаридами, надо соблюдать правила личной гигиены.
|
||
|
64)Цитологические основы универсальности законов Менделя Цитологические основы законов Менделя базируются на: 1) парности хромосом (парности генов, обусловливающих возможность развития какого-либо признака) 2) особенностях мейоза (процессах, происходящих в мейозе, которые обеспечивают независимое расхождение хромосом с находящимися на них генами к разным полюсам клетки, а затем и в разные гаметы) 3) особенностях процесса оплодотворения (случайного комбинирования хромосом, несущих по одному из каждой аллельной пары)
|
20) Понятие об окислительном фосфорилировании. Окисление на уровне ЭТЦ Окислительное фосфорилирование –метаболический путь,при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях в виде АТФ. Этц-служит для передачи электронов от восстановителя субстратов на кислород,что сопровождается трансмембранным переносом ионов Н+ .Таким образом , ЭТЦ митохондрий выполняет функцию ок-вос.Н-помпы (ЭТЦ локализированная во внутренней мембране митохондрий)
|
24)Фотолиз воды: образующиеся продукты ФОТОЛИЗ ВОДЫ — расщепление молекулы воды, в частности в процессе фотосинтеза. Вследствие фотолиза воды образуются протоны (Н+), электроны (е) и свободный кислород, выделяющийся зелеными растениями на свету. Уравнение фотолиза воды 2Н2О (свет) => 4H(+) + 4e(-) + O2 Электроны, образующиеся при фотолизе, восполняют потери их хлорофиллом (как говорят, заполняют «дырку» , возникшую в хлорофилле) . Часть электронов при участии протонов восстанавливает НАДФ+ до НАДФ*Н. Кислород - побочный продукт этой реакции
|
||
|
65)Аллельные гены.. Определение. Формы: взаимодействия,множественный аллелизм,его примеры. Аллельные гены - гены, определяющие альтернативное развитие одного и того же признака и расположенные в идентичных участках гомологичных хромосом. Итак, гетерозиготные особи имеют в каждой клетке два гена - А и а, отвечающих за развитие одного и того же признака. Такие парные гены называют аллельными генами или аллелями. Любой диплоидный организм, будь то растение, животное или человек, содержит в каждой клетке два аллеля любого гена. Исключение составляют половые клетки - гаметы. В результате мейоза в каждой гамете остается один комплект гомологичных хромосом, поэтому любая гамета имеет лишь по одному аллельному гену. Аллели одного гена располагаются в одном и том же месте гомологичных хромосом. Схематически гетерозиготная особь обозначается так: А/а. Гомозиготные особи при подобном обозначении выглядят так: А/А или а/а, но их можно записать и как АА и аа.
Взаимодействие между аллельными генами осуществляется в виде трех форм: полное доминирование, неполное доминирование и независимое проявление (кодоминирование). Полное доминирование – когда один доминантный аллель полностью подавляет проявление рецессивного аллеля, например, желтая окраска горошин доминирует над зеленой. Неполное доминирование наблюдается в том случае, когда один ген из пары аллелей не обеспечивает образование в достаточном для нормального проявления признака его белкового продукта. При этой форме взаимодействия генов все гетерозиготы и гомозиготы значительно отличаются по фенотипу друг от друга. Примером расщепления при неполном доминировании может служить наследование окраски цветков Ночной красавицы. При скрещивании растений с красными цветками (АА) и растений с белыми (аа) гибриды F1 имеют розовые цветки (Аа). Таким образом, имеет место неполное доминирование; в F2 наблюдается расщепление 1 : 2 : 1 как по фенотипу, так и по генотипу. Кроме полного и неполного доминирования известны случаи отсутствия доминантно-рецессивных отношений или кодоминирования. При кодоминировании у гетерозиготных организмов каждый из аллельных генов вызывает формирование в фенотипе контролируемого им признака. Примером этой формы взаимодействия аллелей служит наследование групп крови человека по системе АВ0, детерминируемых геном I. Существует три аллеля этого гена Io, Ia, Ib, определяющие антигены групп крови. Наследование групп крови иллюстрирует также явление множественного аллелизма: в генофондах популяций человека ген I существует в виде трех разных аллелей, которые комбинируются у отдельных индивидуумов только попарно. До этого примера мы говорили о генах, существующих только в двух разных аллельных формах. Однако многие гены состоят из сотен пар нуклеотидов, так что мутации могут проходить во многих участках гена и порождать множество различных его аллельных форм. Так как в каждой из гомологичной хромосом имеется по одному аллельному гену, то, разумеется, диплоидный организм имеет не более двух из серии аллелей генофонда популяции.
|
82)Значение генетики для медицины. Методы изучения наследственности человека Медицинская генетика – раздел генетики, изучающий наследственные заболевания, методы их диагностики и лечения, механизмы их наследования. Методы общей и медицинской генетики — методы, при помощи которых возможно определять закономерности наследственности и изменчивости. Все методы связаны с тем, что изучение наследования признаков человека имеет ряд трудностей: - достаточно сложный кариотип - малое число потомков - позднее половое созревание - редкая смена поколений - невозможность экспериментальных исследований или их затрудненность В генетике используются следующие основные методы: генеалогический (родословный), близнецовый, популяционно-статистический, биохимический, цитогенетический. 1. Клинико-генеалогический метод – 1883 г., Ф.Гальтон. Основан на построении родословной. При этом можно проследить передачу признака в ряду поколений. Позволяет определить: - является ли какой-либо исследуемый признак наследственным - тип и характер наследования признаков (аутосомное и сцепленное с полом наследование, неполное доминирование), (Хсц: ХА, Ха; Усц – голандрическое наследование) - вероятность рождения в семье ребенка с наследственной патологией (генетический риск) - пенетрантность данного признака – частота проявления данного гена у всех лиц его несущих; может быть полная (100%) и неполная (0-100%). Пробанд – лицо, с которого начинается исследование семьи. 2. Близнецовый метод – введен Ф.Гальтоном в 1876 г. Используется для выяснения степени наследственной обусловленности исследуемых признаков; объект исследования – близнецы. Можно изучить степень влияния условий среды на экспрессию генов близнецов. Экспрессивность гена – степень проявления признака, контролируемая данным геном. 3. Популяционно-статистический метод – основан на закономерности Харди-Вайнберга, 1908 г. Позволяет определить частоты генов и фенотипов в достаточно крупных популяциях людей (в разных странах), дает возможность анализа распространения конкретных признаков. Уравнение Харди-Вайнберга: p2+2pq+q2=1 p2 – AA q2 – aa 2pq – Aa 4. Биохимические методы – связаны с изучением заболеваний, в основе которых лежат генные мутации, что приводит к нарушению биосинтеза белка. Заболевание: ферментопатии – связаны с нарушением работы каких-либо ферментов. Можно определить с помощью исследования, идущего с использованием биологических жидкостей. 5. Цитогенетический метод – используют как для нормального кариотипа человека, так и для диагностики заболеваний, связанных с геномными и хромосомными мутациями. Можно понять какой патологией обладает человек. |
72) Геномные мутации Главная отличительная черта геномных мутаций связана с нарушением числа хромосом в кариотипе. Эти мутации так же подразделяются на два вида: полиплоидные и анеуплоидные. Полиплоидные мутации ведут к изменению хромосом в кариотипе, которое кратно гаплоидному набору хромосом. Причины: двойное оплодотворение и отсутствие первого мейотического деления. У человека полиплоидия, а также большинство анеуплоидий приводят к формированию леталей. Этот синдром впервые был лишь обнаружен в 60-ых годах. Вообще полиплодия характерна в основном для человека, а среди животных встречается крайне редко. При полиплоидии число хромосом в клетке насчитывается по 69 (триплодие), а иногда и по 92 (тетраплодие) хромосомы. Такое изменение ведет практически к 100 % смерти зародыша. Триплодие имеет не только многочисленные пороки, но и приводит к потере жизнеспособности. Тетраплодие встречается еще реже, но так же зачастую приводит к летальному исходу. Анеуплоидия — изменение (уменьшение — моносомия, увеличение — трисомия) числа хромосом в диплоидном наборе, т.е. не кратное гаплоидному (2n+1, 2n-1 и т.д.). Механизмы возникновения: нерасхождение хромосом (хромосомы в анафазе отходят к одному полюсу, при этом на каждую гамету с одной лишней хромосомой приходится другая — без одной хромосомы) и «анафазное отставание» (в анафазе одна из передвигаемых хромосом отстаёт от всех других). В результате такой мутации возникают особи с аномальным числом хромосом. Как и триплодия, анеуплодия часто приводит к смерти еще на ранних этапах развития зародыша. Причиной же таких последствий является утрата целой группы сцепления генов в кариотипе. Трисомия — наличие трёх гомологичных хромосом в кариотипе (например, по 21-й паре, что приводит к развитию синдрома Дауна; по 18-й паре — синдрома Эдвардса; по 13-й паре — синдрома Патау). Моносомия — наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по хромосоме X — приводит к развитию синдрома Шерешевского—Тернера (45,Х0).
В цело же, механизм возникновения геномных мутаций связан с патологией нарушения нормального расхождения хромосом в мейозе, в результате чего образуются аномальные гаметы, что и ведет к мутации. Изменения в организме связаны с присутствием генетически разнородных клеток. Такой процесс называется мозаицизм. Геномные мутации одни из самых страшных. Они ведут к таким заболеваниям, как синдром Дауна (трисомия, возникает с частотой 1 больной на 600 новорожденных), синдром Клайнфельтера и др.
|
||
|
31)Митохондрии как энергетические органеллы Митохондрия состоит из наружной мембраны, которая является ее оболочкой и внутренней мембраны, места энергетических преобразований. Внутренняя мембрана образует многочисленные складки, способствующие интенсивной деятельности по преобразованию энергии. Митохондрия - это единственный источник энергии клеток. Расположенные в цитоплазме каждой клетки, митохондрии сравнимы с «батарейками», которые производят, хранят и распределяют необходимую для клетки энергию. Митохондрии подвижны и перемещаются в цитоплазме в зависимости от потребностей клетки. Благодаря наличию собственной ДНК они размножаются и самоуничтожаются независимо от деления клетки. Клетки не могут функционировать без митохондрий, без них жизнь не возможна. СпецифическаяДНК: Самая примечательная особенность митохондрий - это наличие у них своей собственной ДНК: митохондриальной ДНК. Независимо от ядерной ДНК, каждая митохондрия имеет свой собственный генетический аппарат. Митохондрия: энергетическая станция клетки Процесс преобразования клеточной энергии Митохондрии используют 80% кислорода, который мы вдыхаем, чтобы преобразовывать потенциальную энергию в энергию, используемую клеткой. В процессе окисления освобождается большое количество энергии, которая сохраняется митохондриями в виде молекул АТФ. В день преобразовывается 40 кг. АТФ. Энергия в клетке может принимать различные формы. Принцип действия клеточного механизма – преобразование потенциальной энергии в энергию, которую может напрямую использовать клетка. Потенциальные виды энергии попадают в клетку через питание в виде углеводов, жиров и белков Клеточная энергия состоит из молекулы называемой АТФ: Аденозинтрифосфат. Она синтезируется в результате преобразования углеводов, жиров и белков внутри митохондрии. За день в организме взрослого человека синтезируется и распадается эквивалент 40 кг АТФ. |
70) Онтогенетическая изменчивость Онтогенетическая изменчивость– закономерное изменение генотипа и фенотипа в ходе онтогенеза. Изменение фенотипа организма человека в процессе роста, появление вторичных половых признаков – это примеры онтогенетической изменчивости. Закономерное изменение генотипа в ходе онтогенеза обнаружено недавно. Правда, известно таких примеров немного. Так, белки иммуноглобулины у мышей состоят из двух фракций: V (вариабельная) и С (константная). У эмбрионов мышей кодирующие их гены расположены на довольно большом расстоянии друг от друга: Онтогенетическая изменчивость — изменчивость, происходящая в процессе жизни организма и представляющая собой различие между молодым и взрослым организмами на разных этапах развития (напр., молодые растения часто имеют более простое строение листовой пластинки, которая в процессе роста растения усложняется). Она является разновидностью фенотипической изменчивости, которая связана с определенной схемой развития организма в процессе онтогенеза, при этом генотип не претерпевает изменений, а фенотип меняется в соответствии с каждым этапом развития, благодаря морфогенезу и дифференцировке клеток. Морфогенез — это возникновение новых структур на каждом этапе развития, определяемое генетическим аппаратом клеток, может осуществляться благодаря контактным и дистантным межклеточным взаимодействиям, которые контролируют этот процесс. В случае нарушений морфогенеза возникают тератомы (уродства), в том числе и новообразования. Поскольку эти механизмы связаны с «включением» и «выключением» генов, изменчивость этого рода называется — «парагеномная», «эпигенетическая», «эпигенотипическая» или «эпигеномная». |
55)Цикл развития кошачьего сосальщика,пути заражения. Лабораторная диагностика и профилактика описторхоза Кошачий сосальщик (Opisthorchis felineiis) паразитирует в желчных протоках печени, желчном пузыре, поджелудочной железе у человека и многих млекопитающих (кошки, собаки, свиньи и др.) и вызывает заболевание - описторхоз. Распространено это заболевание в бассейнах Оби, Иртыша, Камы, Днепра. Описторхоз - природно-очаговое заболевание. В природе очаги поддерживают животные, питающиеся рыбой. Заражение описторхозом окончательных хозяев, в том числе и человека, происходит при употреблении в пищу сырой, малосоленой, вяленой рыбы из семейства карповых . В мышцах рыб находятся инвазионные стадии паразита - метацеркарии. После попадания в кишечник окончательного хозяина метацеркарии проникают в печень, желчный пузырь, где превращаются во взрослых гельминтов. Взрослый кошачий сосальщик имеет 8 - 12 мм в длину и 1,2 - 2 мм в ширину. Передний конец уже заднего. Есть ротовая и брюшная присоски. Имеются пищеварительная, выделительная, половая и нервная системы. Кишечник не разветвлен, слепо замкнут. Матка расположена в передней части тела, в задней - видны два семенника. В цикле развития кошачьего сосальщика два промежуточных хозяина: первый - пресноводный моллюск; второй - рыбы из семейства карповых. Яйца с фекалиями попадают в воду и заглатываются моллюсками. Из яиц выходят мирацидии, далее образуются спороцисты, редии и церкарий, которые выходят из моллюска и способны самостоятельно плавать. Церкарии проникают во второго промежуточного хозяина - рыбу - и превращаются в метацеркарии. Диагноз описторхоза ставят при нахождении яиц в фекалиях. Профилактика связана с охраной водоемов от загрязнения сточными водами. Кроме того, необходимо обрабатывать термически рыбу перед употреблением. |
||
|
25)Хлорофилл, особенности строения и расположения в хлоропластах Хлорофилл—зеленый пигмент листьев—состоит нз двух близких по строению веществ хлорофилла а—сине-зеленого и хлорофилла Ь—желто-зеленого цвета. Хлорофилл можно назвать "зеленой кровью" мира растений. В растительном мире он выполняет роль протеина. Если сравнить его с молекулой гемоглобина, выполняющей роль переносчика кислорода в крови человека, то можно сказать, что хлорофилл почти идентичен гемоглобину. Единственное отличие заключается в том, что в центре хелатного комплекса в хлорофилле находится атом магния, а в гемоглобине -железо. Поэтому хлорофилл способен оказывать на кровь воздействие сходное с действием гемоглобина: повышать уровень кислорода, ускорять азотистый обмен. Хлорофилл расположен в мембранах тилакоидов гран между слоями молекул белков и липидов.(Хлоропласт- имеет двухмембранное строение,наружная мембрана-гладкая,внутренняя образует систему двухслойных пластин-тилакоидов стромы и тилакоидов гран)
|
26)Биохимическая фаза фотосинтеза. Цикл Кальвина-Бенсона Осуществляется в строме хлоропласта,куда поступают АТФ, НАДФ*Н2 от тилакоидов гран и СО2 из воздуха. Кроме того, там постоянно находятся пятиуглеродные соединения-пентозы С5, которые обазуются в цикле Кальвина(цикле фиксации СО2) Упрощенно этот процесс: 1)К пентозе С5 присоединяется СО2, в результате чего появляется нестойкое шестиуглеродное соединение С6,которое расщепляется на две трехуглеродные группы 2С3-триозы 2)Каждая из триоз 2С3 принимает по одной фосфатной группе от двух АТФ, что обогащает молекулы энергией. 3)Каждая из триоз 2С3 присоединяет по одному атому водорода от двух НАДФ*Н2 4)После чего одни триозы обЬединяются , образуя углеводы 2С3С6С6Н12О6 5)Другие триозы обЬединяются , образуя пентозы 5С33С5, и вновь включаются в цикл фиксации СО2. Суммарная реакция фотосинтеза: 6СО2+6Н2О(энергия светта,хлорофилл) С6Н12О6+6О2 |
63)Законы жизни: Стабильность и изменчивость как важнейшие свойства генетического материала ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ - компоненты клетки, структурно-функциональное единство которых обеспечивает хранение, реализацию и передачу наследственной информации при вегетативном и половом размножении. Генетический материал обладает универсальными свойствами живого: дискретностью, непрерывностью, линейностью, относительной стабильностью генетическая стабильность — Поддерживаемая отбором способность адаптированных ко внешним условиям особей воспроизводить потомство с такой же генетической конституцией.
Генетическая стабильность популяций. Равновесие Харди - Вайнберга (1908). Генетическая стабильность популяции выражается в формулировке закона Харди-Вайнберга, который отражает равновесие частот генов и генотипов в панмиктической популяции: «В безгранично большой популяции в отсутствии мутаций, избирательной миграции организмов и давления естественного отбора первоначальная частота аллелей (генов и генотипов) остается неизменной из поколения в поколение». ИЗМЕНЧИВОСТЬ - это способность дочерних организмов отличаться от родительских форм морфологическими и физиологическими признаками и особенностями индивидуального развития. |
||
|
4)Клеточная теория: история,современное состояние,значение для биологии и медицины. Клеточная теория позволила сформулировать вывод о том, что клетка – это важнейшая составляющая часть всех живых организмов. Клеточная теория позволила прийти к выводу о сходстве химического состава всех клеток и еще раз подтвердила единство всего органического мира. Современная клеточная теория включает следующие положения. 1.Клетка – основная единица строения и развития всех живых организмов, наименьшая единица живого. 2. Клетки всех одноклеточных и многоклеточных организмов сходны (гомологичны) по своему строению, химическому составу, основным проявлениям жизнедеятельности и обмену веществ. 3. Размножение клеток происходит путем их деления, и каждая новая клетка образуется в результате деления материнской клетки. 4. В сложных многоклеточных организмах клетки специализированы по выполняемым ими функциям и образуют ткани; из тканей состоят органы, которые тесно взаимосвязаны и подчинены нервным и гуморальным системам регуляции. 5.Клеточное строение всех ныне живущих организмов-свидетельство единства происхождения Значение клеточной теории Клеточная теория позволила понять как зарождается, развивается и функционирует живой организм, то есть создала основу эволюционной теории развития жизни, а в медицине – понимания процессов жизнедеятельности и развития болезней на клеточном уровне – что открыло немыслимые ранее новые возможности диагностики, лечение заболеваний. Cтало ясно, что клетка — важнейшая составляющая часть живых организмов, их главный морфофизиологический компонент. Клетка — это основа многоклеточного организма, место протекания биохимических и физиологических процессов в организме. На клеточном уровне в конечном итоге происходят все биологические процессы. Клеточная теория позволила сделать вывод о сходстве химического состава всех клеток, общем плане их строения, что подтверждает филогенетическое единство всего живого мира. |
43)Эмбриогенез: образование тканей и органов. ГИСТО- И ОРГАНОГЕНЕЗ После образования мезодермы начинается процесс гисто- и органогенеза. Сначала формируются осевые органы - нервная трубка, хорда, затем все остальные. У ланцетника из эктодермы на спинной стороне зародыша образуется нервная трубка .Остальная эктодерма формирует кожный эпителий и его производные. Из энто- и мезодермы под нервной трубкой формируется хорда. Под хордой находится кишечная трубка энтодермального происхождения. По бокам от хорды - мезодерма сомитов, которая дифференцирована следующим образом: наружная часть сомита, прилегающая к эктодерме, называется дермотомом. Из нее образуется соединительная ткань кожи. Внутренняя часть - склеротом - дает начало скелету. Между дермотомом и склеротомом находится миотом, дающий начало поперечно-полосатой мускулатуре. Под сомитами расположены его ножки (нефрогонотом), из которых формируется мочеполовая система. Целомические мешки образуются симметрично по бокам. Внутри целомических мешков находится вторичная полость тела - целом, - заполненная жидкостью. Стенки целомических мешков, обращенные в сторону кишечника, называют спланхноплеврой, в сторону эктодермы - соматоплеврой. Эти листки участвуют в образовании сердечнососудистой системы, плевры, брюшины, перикарда. первичной кишки (гастроцель), знаком "х" - полость кишечной трубки, в которую замкнулась кишечная энтодерма. Таким образом, из эктодермы формируются наружный эпителий, кожные железы, эмаль зубов, волосы, ногти, когти. Из энтодермы образуются эпителий средней кишки, печень, поджелудочная железа, щитовидная железа, тимус, эпителий дыхательной системы. Мезодерма участвует в образовании мышц, соединительной ткани, костной ткани, каналов выделительной системы, кровеносной системы, части ткани половых желез. |
18)Понятие об ЭТЦ и дыхательной цепи митохондрий Дыхательная цепь переноса электронов (ЭТЦ)— система структурно и функционально связанных трансмембранных белков и переносчиков электронов. ЭТЦ запасает энергию, выделяющуюся в ходе окисления НАД∙Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану. Дыхательная цепь– это совокупностьred-oxреакций в результате которых Н2 и электроны переносятся на О2 с выделением Н2О и энергии. В результате окислительных реакций в цикле Кребса, катализируемых дегидрогеназами, протоны и электроны переходят от органических субстратов на кофакторы НАД и ФАД, восстанавливая их в НАДН иФАДН2. Эти вещества обладают высоким энергетическим потенциалом. Электроны от них передаются на О2 ч/з ряд red-ox реакций. Окисление органических веществ в клетках сопровождающееся потреблением О2 и образованием Н2О называется тканевым дыханием, а цепь переноса электронов – дыхательной цепью. Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных кислот, спиртов и АК, далее поступают в митохондрии, где ферментами дыхательной цепи осуществляется процесс окислительного фосфорилирования. Дыхательная цепь является частью процесса окислительного фосфорилирования. Компоненты дыхательной цепи катализируют перенос электронов от НАДН + Н+ или восстановленного убихинона (QH2) на молекулярный кислород. Из-за большой разности окислительно-восстановительных потенциалов донора (НАДН + Н+ и, соответственно, (QH2) и акцептора (О2) реакция является высокоэкзергонической. Большая часть выделяющейся при этом энергии используется для создания градиента протонов и, наконец, для образования АТФ с помощью АТФ-синтазы |
||
|
|
|
|
||
|
71)Генные мутации, молекулярные механизмы, определяющие частоту мутаций в природе. ГЕННЫЕ МУТАЦИИ связаны с изменением состава или последовательности нуклеотидов ДНК в пределах гена. Генные мутации наиболее важны среди всех категорий мутаций. Синтез белка основан на соответствии расположения нуклеотидов в гене и порядком аминокислот в молекуле белка. Возникновение генных мутаций (изменение состава и последовательности нуклеотидов) изменяет состав соответствующих белков-ферментов и в итоге к фенотипическим изменениям. Мутации могут затрагивать все особенности морфологии, физиологии и биохимии организмов. Многие наследственные болезни человека также обусловлены мутациями генов. Мутации в естественных условиях случаются редко - одна мутация определенного гена на 1000-100000 клеток. Но мутационный процесс идет постоянно, идет постоянное накопление мутаций в генотипах. А если учесть, что число генов в организме велико, то можно сказать, что в генотипах всех живых организмов имеется значительное число генных мутаций.
По своим последствиям генные мутации делятся на две группы: мутации без сдвига рамки считывания и мутации со сдвигом рамки считывания. Мутации без сдвига рамки считывания происходят в результате замены нуклеотидных пар, при этом общая длина ДНК не изменяется. В результате возможна замена аминокислот, однако из-за вырожденности генетического кода возможно и сохранение структуры белка. Мутации со сдвигом рамки считывания (фреймшифты) происходят в результате вставки или потери нуклеотидных пар, при этом общая длина ДНК изменяется. В результате происходит полное изменение структуры белка. Однако если после вставки пары нуклеотидов происходит потеря пары нуклеотидов (или наоборот), то аминокислотный состав белков может восстановиться. Тогда две мутации хотя бы частично компенсируют друг друга. Это явление называется внутригенной супрессией. |
51)Цикл развития малярийного плазмодия Человек-промежуточный хозяин бесполая фаза паразита:Человек инфицируется спорозоитами(при укусе малярийного комара) 1)Скрытый период болезни:спорозоиты паразитируют и размножаются путем шизогонии в сосудах и печени;из спорозоитов образуются мерозоиты; 2)мерозоиты внедряются в эритроциты(цикл повторяется каждые три дня);мерозоиты образуют гаметоциты Самка малярийного комара-основной хозяин Половая фаза: гаметоциты попадают в желудок малярийного комара(при укусе больного чпеловека); из гаметоцитов образуются яйцеклетки и сперматозоиды;зиготова(оокинета);ооциста;мейоз;митоз спорозоиты;спорозоиты попадают в слюну самки комара;человек инфицируется спорозоитами(при укусу малярийного комара,анофелесом)…(Мейоз-в стенке кишечника)
Со слюной зараженного комара при укусе плазмодии попадают в кровь человека. С током крови они разносятся по организму и поселяются в клетках печени. Здесь они растут и размножаются. Клетки печени при этом разрушаются и паразиты, называющиеся на этой стадии мерозоитами, поступают в кровь и внедряются в эритроциты. С этого момента начинается эритроцитарная часть цикла развития плазмодия. Паразит питается гемоглобином, растет и размножается шизогонией. Следующая стадия называетсяамебовидным шизонтом. У паразита появляются ложноножки, а вакуоль увеличивается. Наконец плазмодий занимает почти весь эритроцит. Следующая стадия развития паразита —фрагментация шизонта. После разрушения эритроцита мерозоиты попадают в плазму крови и оттуда в новые эритроциты, после чего весь цикл эритроцитарной шизогонии повторяется. Таким образом, в организме человека плазмодий размножается только бесполым путем — шизогонией, человек является его промежуточным хозяином. В организме комара проходят две другие стадии цикла развития п аразита: половой процесс — гаметогония и образование спорозоитов за счет деления под оболочкой ооцисты — спорогония. Поэтому малярийный комар является окончательным хозяином этого паразита.
|
79)Репарационные системы(понятие,пример) Два типа нарушений структуры ДНК приводят к мутациям. Это, во-первых, включение нормальных нуклеотидов в аномальное окружение из последовательностей нуклеотидов, приводящих к образованию неправильно спаренных оснований и петель разных размеров. Во-вторых, появление повреждений ДНК в виде аномальных нуклеотидов в правильных последовательностях ДНК. В этом случае речь идет о различных химических модификациях нуклеотидов, включая их разрушение и образование поперечных сшивок. Повреждения ДНК могут приводить к задержке и блокированию репликации и транскрипции.
При исследовании механизмов репарации ДНК важные результаты были получены на клетках, облученных УФ-светом с длинами волн 240-280 нм. УФ-облучение клеток часто сопровождается их гибелью, образованием мутаций и злокачественной трансформацией. Среди первичных повреждений наиболее часто встречаются биспиримидиновые фотопродукты: пиримидиновые димеры циклобутанового типа, соединенные связью 6-4 ( рис. I.56 ). Как про-, так и эукариоты имеют несколько ферментных систем, которые разделяют пиримидиновые димеры или восстанавливают исходную структуру азотистых оснований. К таким репаративным системам относится, прежде всего, система эксцизионной репарации ДНК (NER) , осуществляющая вырезание поврежденных нуклеотидов ( NER - nucleotide excision repair ) или азотистых оснований ( BER - base excision repair ). Система ферментативной фотореактивации ДНК ( PHR - photoreactivation ), основным компонентом которой является ДНК- фотолиаза, разделяет пиримидиновые димеры, превращая их в нормальные пиримидиновые основания. Кроме того, поврежденные УФ- светом молекулы ДНК могут репарироваться с участием систем рекомбинации и в процессе пострепликативного синтеза ДНК. Действие систем репарации поврежденной ДНК распространяется не только на фотопродукты, но и на другие модифицированные основания, образующиеся под действием химических мутагенов. Отдельно следует упомянуть систему, распознающую неправильно спаренные основания в двойной спирали ДНК, возникающие в результате ошибок репликации |
||
|
12)Ассимиляция и диссимиляция как основа самообновления биологических систем. Определение, сущность, значение. Основные реакции ассимиляции. Все живые организмы способны к обмену веществ с окружающей средой, поглощая из нее элементы, необходимые для питания, и выделяя продукты жизнедеятельности. В круговороте органических веществ самыми существенными стали процессы синтеза и распада. Живые организмы поглощают из окружающей среды различные вещества. Вследствие целого ряда сложных химических превращений вещества из окружающей среды уподобляются веществам живого организма, из них строится его тело. Эти процессы называются ассимиляцией или пластическим обменом. Приведем несколько примеров. Растения из диоксида углерода и воды синтезируют сложные органические соединения – углеводы (крахмал и целлюлозу), которые используются как запасные питательные вещества и строительный материал. Белок куриного яйца в организме человека претерпевает ряд сложных превращений, прежде чем преобразуется в белки, свойственные организму, - гемоглобин, кератин или любой иной. Другая сторона обмена веществ – процессы диссимиляции, в результате которых сложные органические соединения распадаются на простые соединения, при этом утрачивается их сходство с веществами организма и выделяется энергия, необходимая для реакций биосинтеза. Поэтому диссимиляцию называют еще энергетическим обменом. Обмен веществ обеспечивает постоянство химического состава и строения всех частей организма и как следствие – постоянство функционирования в непрерывно меняющихся условиях окружающей среды.
|
|
|||
|
16)Механизмы переноса веществ через мембраны митохондрий Одна из главных функций мембран – регуляция переноса веществ в клетку и из клетки, сохранение веществ, которые нужны клетке, и освобождение от ненужных. Прохождение веществ через мембраны может проходить по градиенту концентрации – пассивный транспорт и против градиента концентрации – активный транспорт. Любая молекула может пройти через липидный бислой, однако скорость пассивной диффузии веществ, т.е. перехода вещества из области с большей концентрацией в область с меньшей, может сильно отличаться. Для некоторых молекул это занимает столь длительное время, что можно говорить об их практической непроницаемости для липидного бислоя мембраны. Скорость диффузии веществ через мембрану зависит главным образом от размера молекул и их относительной растворимости в жирах. Легче всего проходят простой диффузией через липидную мембрану малые неполярные молекулы, такие как О2, стероиды, а также жирные кислоты. Малые полярные незаряженные молекулы - СО2, NH3, Н2О, этанол, мочевина - также диффундируют с достаточно большой скоростью. Диффузия глицерола идёт значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана непроницаема. Транспорт таких молекул возможен благодаря наличию в мембранах либо белков, формирующих в липидном слое каналы (поры), заполненные водой, через которые могут проходить вещества определённого размера простой диффузией, либо специфических белков-переносчиков, которые избирательно взаимодействуя с определёнными лигандами, облегчают их перенос через мембрану (облегчённая диффузия). Облегчённая диффузия веществ В мембранах клеток существуют белки-транслоказы. Взаимодействуя со специфическим лигандом, они обеспечивают его диффузию (транспорт из области большей концентрации в область меньшей) через мембрану. В отличие от белковых каналов, транслоказы в процессе взаимодействия с лигандом и переноса его через мембрану претерпевают конформационные изменения. Кинетически перенос веществ облегчённой диффузией напоминает ферментативную реакцию. Для транслоказ существует насыщающая концентрация лиганда, при которой все центры связывания белка с лигандом заняты, и белки работают с максимальной скоростью Vmax. Поэтому скорость транспорта веществ облегчённой диффузией зависит не только от градиента концентраций переносимого лиганда, но и от количества белков-переносчиков в мембране. Существуют транслоказы, переносящие только одно растворимое в воде вещество с одной стороны мембраны на другую. Такой простой транспорт называют "пассивный унипорт". Примером унипорта может служить функционирование ГЛЮТ-1 - транслоказы, переносящей глюкозу через мембрану эритроцита Облегчённая диффузия (унипорт) глюкозы в эритроциты с помощью ГЛЮТ-1 (S - молекула глюкозы). Молекула глюкозы связывается переносчиком на наружной поверхности плазматической мембраны. Происходит конформационное изменение, и центр переносчика, занятый глюкозой, оказывается открытым внутрь клетки. Вследствие конформационных изменений переносчик теряет сродство к глюкозе, и молекула высвобождается в цитозоль клетки. Отделение глюкозы от переносчика вызывает конформационные изменения белка, и он возвращается к исходной "информации. Некоторые транслоказы могут переносить два разных вещества по градиенту концентраций в одном направлении - пассивный симпорт, или в противоположных направлениях - пассивный антипорт. Типы (виды) облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные молекулы. Примером транслоказы, работающей по механизму пассивного антипорта, может служить анионный переносчик мембраны эритроцитов. Пассивный антипорт анионов НСО3- и Сl- через мембрану эритроцитов. А - когда эритроцит находится в венозных капиллярах, анион НСО3-, образованный при диссоциации угольной кислоты, по градиенту концентрации выходит в кровь. В обмен на каждый транспортируемый из клетки ион НСО3- транслоказа переносит в эритроцит ион Cl-; Б - когда кровь достигает лёгких транслоказа производит обмен ионами в противоположных направлениях. Такая "челночная" система работает очень быстро и обеспечивает удаление СО2 из организма и в то же время сохранение оптимального значения рН в клетке. Внутренняя митохондриальная мембрана содержит много транслоказ, осуществляющих пассивный антипорт.
|
14)Гликолиз. Окисление на уровне субстрата На первой стадии Г.происходит фосфорилирование молекулы глюкозы (при этом расходуется энергия АТФ) и расщепление гексозы с образованием двух взаимопревращающихся триозофосфатов — диоксиацетонфосфата и глицеральдегид-3-фосфата. На первой стадии в гликолиз вовлекаются и другие простые сахара (как правило, путем превращения в глюкозо-6-фосфат), а также полисахариды. Образование фосфорных производных сахаров способствует превращению циклических форм сахаров в более реакционно-способные формы — ациклические. Гликолитическое расщепление гликогена, осуществляемое главным образом в мышцах и печени, называется гликогенолизом. Первой реакцией гликогенолиза является фосфоролиз гликогена при участии фермента гликогенфосфорилазы с образованием глюкозо-1-фосфата, в реакции, катализируемой фосфоглюкомутазой, превращающегося в глюкозо-6-фосфат, который затем подвергается дальнейшим гликолитическим превращениям. У растений субстратом гликолиза может служить крахмал. Вторая стадия гликолиза является общей для всех сахаров, участвующих в Г. , и состоит в превращении глицеральдегид-3-фосфата в лактат. Центральным звеном этой стадии является гликолитическая оксидоредукция, включающая окислительно-восстановительные реакции, сопряженные с аккумуляцией энергии в виде АТФ в процессе фосфорилирования аденозиндифосфорной кислоты (АДФ) на уровне субстрата. Энергия, высвобождающаяся в результате превращения глицеральдегид-3-фосфата в 3-фосфоглицерат запасается в виде макроэргических фосфатных связей АТФ. Образование АТФ происходит также при разрыве макроэргической фосфатной связи фосфоенолпировиноградной кислоты (фосфоенолпирувата) и переносе фосфорильного остатка на АДФ. Внутренний окислительно-восстановительный цикл гликолиза завершается восстановлением пировиноградной кислоты или пирувата (-кетопропионовой кислоты). Эта простейшая кетокислота занимает центральное место в превращении углеводов и участвует в обмене аминокислот в качестве субстрата трансаминирования. В аэробных условиях пируват подвергается окислительному декарбоксилированию при участии пируватдегидрогеназного мультиферментного комплекса с образованием ацетил-кофермента А (ацетил-КоА); это превращение является одной из стадий тканевого дыхания, итогом которой служит включение углеводов в цикл трикарбоновых кислот в качестве энергетического субстрата. Ацетил-КоА участвует также в метаболизме липидов (см. Жировой обмен) и других физиологически важных соединений (например, ацетилхолина). Скорость гликолиза и ее координация со скоростями других метаболических процессов, в первую очередь глюконеогенеза и цикла трикарбоновых кислот, обеспечивается действием разнообразных регуляторных механизмов. Общая скорость гликолиза определяется доступностью субстрата, использованием АТФ и концентрацией ферментов Г. Существенную роль в регуляции скорости гликолиза на уровне ферментов играют приведенные выше три практически необратимые реакции Г. Наиболее важным лимитирующим скорость гликолиза ферментом является фосфофруктокиназа, активность которой ингибируется АТФ, НАДН, лимонной кислотой (цитратом) и жирными кислотами и стимулируется АДФ и АМФ, Активности гексокиназы и пируваткиназы также регулируются (по принципу обратной связи) АДФ, АТФ, промежуточными продуктами гликолиза и цикла трикарбоновых кислот. Основными пунктами контроля глюконеогенеза на уровне ферментов является регуляция реакций, катализируемых пируваткарбоксилазой (активация ацетил-КоА) и фруктозодифосфатазой (ингибирование АМФ и активация АТФ), т.е. глюконеогенез зависит от наличия избытка субстрата дыхания и энергии для синтеза глюкозы. У животных и человека в регуляции Г. принимают участие гормоны. Так, инсулин осуществляет контроль за гликолизом на генетическом уровне, являясь индуктором образования ключевых ферментов Г. (гексокиназы, фосфофруктокиназы, пируваткиназы) и репрессором синтеза ферментов глюконеогенеза. Противоположным действием обладают катехоламины, глюкагон, АКТГ (в печени) и паратгормон (в почках).
|
|||
|
35)Размножение-основное свойство живого. Бесполое размножение, его формы, биологическое значение Размножение в органическом мире. Способность к размножению является одним из важнейших признаков жизни. Эта способность проявляется уже на молекулярном уровне жизни. Вирусы, проникая в клетки других организмов, воспроизводят свою ДНК или РНК и таким образом размножаются. Размножение – это воспроизведение генетически сходных особей данного вида, обеспечивающее непрерывность и преемственность жизни. Различают следующие формы размножения:
Бесполое размножение. Эта форма размножения характерна как для одноклеточных, так и для многоклеточных организмов. Однако наиболее распространено бесполое размножение в царствах Бактерии, Растения и Грибы. В царстве среди животных этим способом размножаются в основном простейшие и кишечнополостные. В бесполой форме размножение осуществляется родительской особью самостоятельно, без обмена наследственной информацией с другими особями. Дочерний организм образуется путем отделения от родительской особи одной или нескольких соматических (телесных) клеток и дальнейшего их размножения посредством митоза. Потомство наследует признаки родителя, являясь в генетическом отношении его точной копией.
Существует несколько способов бесполого размножения:
– Простое деление материнской клетки на две или несколько клеток. Особенно распространено бесполое размножение у бактерий и сине-зеленых водорослей. Единственная клетка этих безъядерных организмов разделяется пополам или сразу на несколько частей. Каждая часть является целостным функциональным организмом. Простым делением размножаются амебы, инфузории, эвглены и другие простейшие. Разделение происходит посредством митоза, поэтому дочерние организмы получают от родительских тот же набор хромосом.
– Вегетативное размножение частями тела характерно для многоклеточных организмов – растений, губок, кишечнополостных, некоторых червей. Растения вегетативно могут размножаться черенками, отводками, корневыми отпрысками и другими частями организма. Этот вид бесполого размножения широко распространен у растений.
– Почкование – Этот тип размножения используют как одноклеточные, так и некоторые многоклеточные организмы: дрожжи (низшие грибы), инфузории, коралловые полипы. Почкование у пресноводных гидр происходит следующим образом. Сначала на стенке гидры образуется вырост, который постепенно удлиняется. На его конце появляются щупальца и ротовое отверстие. Из почки вырастает маленькая гидра, которая отделяется и становится самостоятельным организмом. У других существ почки могут оставаться на теле родителя.
– Митотическое спорообразование. Родоначальницей нового организма может стать специализированная клетка родительского существа — спора. Такой способ размножения характерен для растений и грибов. Размножаются спорами многоклеточные водоросли, мхи, папоротники, хвощи и плауны.
Бесполое размножение обычно обеспечивает увеличение численности генетически однородного потомства, поэтому его часто применяют селекционеры растений для сохранения полезных свойств сорта.
|
39)Партеногенез,его формы и распростаненность в природе. Партеногенез – развитие без оплодотворения. В случае естественного партеногенеза развитие идет на основе цитоплазмы и пронуклеуса яйцеклетки. Особи, формирующиеся из яйцеклетки, имеют либо гаплоидный, либо диплоидный набор хромосом, так как чаще всего в начале дробления срабатывает один из механизмов удвоения числа хромосом. Естественный партеногенез чаще всего случается при незавершенном оплодотворении, т. е. в тех случаях, когда имела место активация яйцеклетки, но ядро сперматозоида не участвовало в оплодотворении. В активированных яйцах используется информация только женского пронуклеуса. Такой вид партеногенеза называют гиногенезом. При искусственном партеногенезе можно удалить женский пронуклеус, и тогда развитие осуществляется только за счет мужских пронуклеусов. Это андрогенез. Потомки наследуют либо только признаки матери при гиногенезе, либо только признаки отца – при андрогенезе. Это указывает на то, что наследственные свойства особи определяются в основном ядром, а не цитоплазмой. Естественный партеногенез явление редкое, и как правило не является единственным способом размножения вида. У пчел, например, он используется как механизм генотипического определения пола: женские особи (рабочие пчелы и царицы) развиваются из оплодотворенных яйцеклеток, а мужские (трутни) – партеногенетически. Существует несколько классификаций партеногенетического размножения. 1)По способу размножения Естественный — нормальный способ размножения некоторых организмов в природе. Искусственный — вызывается экспериментально действием разных раздражителей на неоплодотворённую яйцеклетку, в норме нуждающуюся в оплодотворении. 2)По полноте протекания Рудиментарный (зачаточный) — неоплодотворённые яйцеклетки начинают деление, однако зародышевое развитие прекращается на ранних стадиях. Вместе с тем в некоторых случаях возможно и продолжение развития до конечных стадий (акцидентальный или случайный партеногенез). Полный — развитие яйцеклетки приводит к формированию взрослой особи. Эта разновидность партеногенеза наблюдается во всех типах беспозвоночных и у некоторых позвоночных. 3) По наличию мейоза в цикле развития Амейотический — развивающиеся яйцеклетки не проделывают мейоза и остаются диплоидными. Такой партеногенез (например, у дафний) является разновидностью клонального размножения. Мейотический — яйцеклетки проделывают мейоз (при этом они становятся гаплоидными). Новый организм развивается из гаплоидной яйцеклетки (самцы перепончатокрылых насекомых и коловраток), или яйцеклетка тем или иным способом восстанавливает диплоидность (например, путём эндомитоза или слияния с полярным тельцем) 4)По наличию других форм размножения в цикле развития Облигатный — когда он является единственным способом размножения Циклический — партеногенез закономерно чередуется с другими способами разножения в жизненном цикле (напрмер, у дафний и коловраток). Факультативный — встречающийся в виде исключения или запасного способа размножения у форм, в норме двуполых. 5)В зависимости от пола организма Гиногенез — партеногенез самок Андрогенез — партеногенез самцов Распространенность У животных У членистоногих Способность к партеногенезу у членистоногих имеют тихоходки, тля, балянус, некоторые муравьи и многие другие. Муравьи У муравьёв телитокический партеногенез обнаружен у 8 видов и может быть разделён на 3 основных типа: тип A — самки производят самок и рабочих через телитокию, но рабочие стерильны и самцы отсутствуют ; тип B — рабочие производят рабочих и потенциальных самок через телитокию; тип C — самки производят самок телитокически, а рабочих — обычным половым путём, в то же время, рабочие производят самок через телитокию. Самцы известны для типов B и C. Партеногенез редок у позвоночных и встречается примерно у 70 видов, что составляет 0,1 % всех позвоночных животных. Например, существует несколько видов ящериц, в естественных условиях размножающихся партеногенезом (Даревскиа, комодские вараны). Партеногенетические популяции также найдены и у некоторых видов рыб, земноводных, птиц (в том числе куриц). Случаи однополого размножения пока не известны только среди млекопитающих. Партеногенез у комодских варанов возможен потому, что оогенез сопровождается развитием полоцита (полярного тельца), содержащего удвоенную копию ДНК яйца; полоцит при этом не погибает и выступает в качестве спермы, превращая яйцеклетку в эмбрион.
|
|||
|
66)Множественные аллели и полигенное наследование на примере человека. Взаимодействие неаллельных генов: комплементарность. Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов: комплемментарность; эпистаз; полимерия. Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые. Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3. Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 но фенотипу происходит в соотношении 1:4:6:4:1. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1. Пример: цвет кожи у людей, который зависит от четырёх генов.
|
78)Понятие о заболеваниях, обЬясняемых законами Менделя Основные закономерности наследования признаков, установленные Менделем 1 При скрещивании чистосортных растений все гибриды первого поколения единообразны и характеризуются доминантным вариантом признака. 2 При скрещивании гибридов первого поколения между собой в их потомстве наблюдается расщепление в соотношении – 3 части растений с доминантным вариантом признака : 1 часть растений с рецессивным вариантом. 3 Отдельные признаки наследуются независимо друг от друга. Современные формулировки законов Менделя 1-й закон Менделя – закон единообразия гибридов первого поколения. При скрещивании гомозигот все гибриды первого поколения единообразны по генотипу и фенотипу. Правило чистоты гамет. При гаметогенезе у гетерозигот в каждую из гамет с равной вероятностью переходит один из двух аллелей. 2-й закон Менделя – закон расщепления. При моногибридном скрещивании гетерозигот примерно четвертая часть их потомков обладает рецессивным вариантом признака. 3-й закон Менделя – закон независимого наследования отдельных признаков. Отдельные признаки наследуются независимо друг от друга, если гены, отвечающие за развитие этих признаков, не сцеплены между собой.
Моногибридное скрещивание Моногибридным называется скрещивание, при котором родительские формы отличаются друг от друга по одной паре контрастных, альтернативных признаков. Признак — любая особенность организма, т. е. любое отдельное его качество или свойство, по которому можно различить две особи. У растений это форма венчика (например, симметричный—асимметричный) или его окраска (пурпурный—белый) Совокупность всех признаков организма, начиная с внешних и кончая особенностями строения и функционирования клеток, тканей и органов, называется фенотипом. Этот термин может употребляться и по отношению к одному из альтернативных признаков. Признаки и свойства организма проявляются под контролем наследственных факторов, т. е. генов. Совокупность всех генов организма называют генотипом. Примерами моногибридного скрещивания, проведенного Г. Менделем, могут служить скрещивания гороха с такими хорошо заметными альтернативными признаками, как пурпурные и белые цветки, желтая и зеленая окраска незрелых плодов (бобов), гладкая и морщинистая поверхность семян, желтая и зеленая их окраска и др. Единообразие гибридов первого поколения (первый закон Менделя). При скрещивании гороха с пурпурными и белыми цветками Мендель обнаружил, что у всех гибридных растений первого поколения (F1)цветки оказались пурпурными. При этом белая окраска цветка не проявлялась. Мендель установил также, что все гибриды F1оказались единообразными (однородными) по каждому из семи исследуемых им признаков. Следовательно, у гибридов первого поколения из пары родительских альтернативных признаков проявляется только один, а признак другого родителя как бы исчезает. Явление преобладания у гибридов F1признаков одного из родителей Мендель назвал доминированием, а соответствующий признак — доминантным. Признаки, не проявляющиеся у гибридов F1 он назвал рецессивными.
Поскольку все гибриды первого поколения единообразны, это явление было названо К. Корренсом первым законаом Менделя, или законом единообразия гибридов первого поколения, а также правилом доминирования.
|
|||