

5.5. Взаимодействие генов
Генотип организма (совокупность всех генов особи) определяют как исторически сложившуюся систему взаимодействующих генов. Несмотря на расположение генов в различных хромосомах или различных локусах одной и той же хромосомы гены могут взаимодействовать. Биохимической основой взаимодействия генов является многоэтапность формирования признака и участие в нём различных ферментов, биосинтез каждого из которых контролируется отдельным геном.
Соответственно разделению генов на аллельные и неаллельные выделяют типы взаимодействия аллельных и неаллельных генов.
5.5.1. Взаимодействие аллельных генов
Аллельные гены определяют альтернативные варианты (формы) развития признака и располагаются в идентичных участках гомологичных хромосом (под признаком при этом понимают дискретное свойство, по которому один организм можно отличать от другого).
Первый тип взаимодействия аллельных генов - полное доминирование (доминирование по Менделю) описано Г. Менделем в 1865 году (рис. 62). Для опытов по скрещиванию Мендель выбрал 22 сорта гороха, которые имели чёткие альтернативные различия по семи признакам: семена круглые или угловатые; семядоли жёлтые или зелёные; кожура семян серая или белая; семена выполненные или морщинистые; цветы пазушные или верхушечные; растения карликовые или высокие. В течение ряда лет Мендель исследовал эти сорта, используя самоопыление растений, пока не убедился, что эти сорта представляют собой наследственно чистые формы и в ряду потомств не дают никаких уклонений от стандартных признаков сорта.
Скрестив сорта, отличающиеся по контрастным взаимоисключающим (альтернативным) признакам, Мендель обнаружил, что гибриды, полученные от такого скрещивания, проявляют только один признак из альтернативной пары. Такой признак Г. Мендель назвал доминантным. Среди них оказалась округлая форма семян, жёлтая окраска семядолей (семян), серый
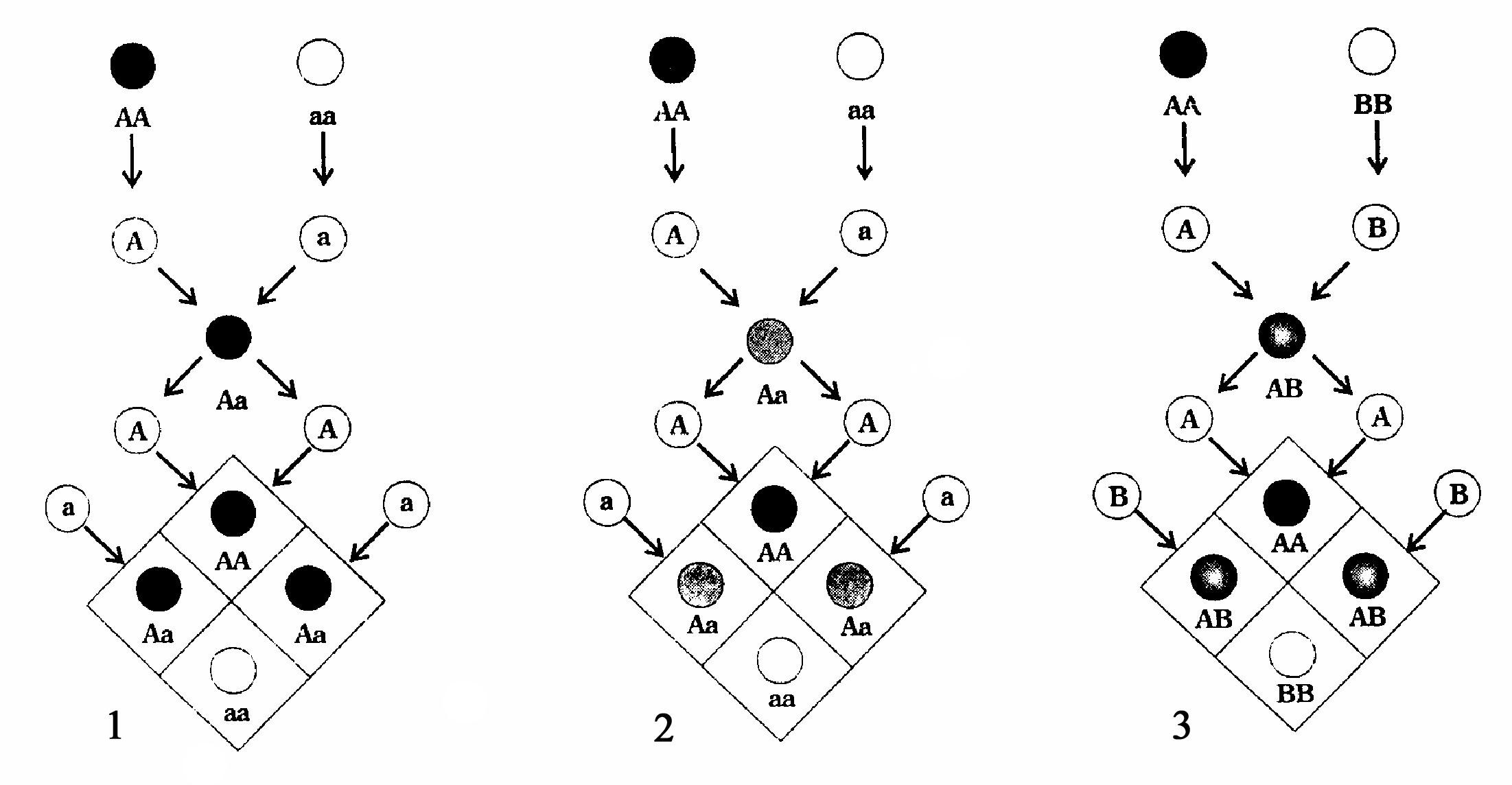
Рис.
62. Схема
взаимодействия аллельных генов:
1 – полное
доминирование; 2 – неполное доминирование;
3 – кодоминирование
цвет семенной кожуры, выполненность боба, зелёная окраска бобов, пазушные цветы, высокий рост растений.
Те факты, что гибриды первого поколения из пары признаков родителей проявляют только один доминантный признак, Мендель обобщил как правило (закон) доминирования. Это правило описывает полное доминирование как определённый тип взаимодействия аллельных генов. Полным доминированием называется тип взаимодействия аллельных генов, при котором один (доминантный) аллельный ген у гибридов первого поколения полностью подавляет проявление другого (рецессивного) аллельного гена.
Для обозначения доминантных (А) и рецессивных (а) аллельных генов У. Бэтсон в 1902 году предложил термин «аллеломорфы». В 1909 году В. Иогансен трансформировал его в термин «аллели». Пара аллелей характеризует два взаимоисключающих состояния гена. Константные формы АА и аа, которые в последующих поколениях не дают расщепления, В. Бэтсон в 1902 году предложил называть гомозиготными, а формы Аа, дающие во втором поколении расщепление по фенотипу в соотношении 3:1, назвал гетерозиготными. Константные признаки, контролируемые разными аллелями генов, обнаружены у всех живых организмов.
Позднее был описан второй тип взаимодействия аллельных генов - неполное (частичное) доминирование (рис. 62). При неполном доминировании гетерозиготная особь имеет фенотип, промежуточный между фенотипами гомозиготных особей. В связи с этим неполное доминирование часто называют промежуточным наследованием. На него распространяется правило Менделя о единообразии фенотипа в первом (F1) поколении. Во втором (F2) поколении расщепление как по фенотипу, так и по генотипу выражается отношением 1:2:1. Примером неполного доминирования может служить промежуточная розовая окраска цветка у гибридов ночной красавицы Mirabilis jalara, полученных от скрещивания красноцветковой и белоцветковой форм. Неполное доминирование оказалось широко распространённым явлением и было отмечено при изучении наследования окраски цветка у львиного зева, окраски оперения у андалузских кур, шерсти у крупного рогатого скота и овец и др.
А
Рис.
63. Выявление
аллелей белков-ферментов с разной
электрофоретической подвижностью: S
(slow)
- медленная и F
(fast)
- быстрая у гибридов F1
 ллельные
гены могут взаимодействовать по типусовместного доминирования
или кодоминирования
(рис. 62). Кодоминирование
- это тип взаимодействия аллельных
генов, при котором оба аллеля проявляют
активность, внося равноценный вклад в
формирование феноти-
па.
Известно, например, что серповидноклеточная
анемия проявляется как аутомно-рецессивное
заболевание в гомозиготном организме.
В таком организме присутствуют два
патологических аллеля одного и того же
гена, контролирующие синтез дефектного
гемоглобина. В гетерозиготном организме
присутствуют нормальный и дефектный
аллельные гены. Причём ни один из них
не доминирует над другим, поэтому в
организме одновременно синтезируются
оба вида гемоглобина (нормальный и
дефектный). У таких индивидов симптоматика
заболевания почти отсутствует или она
проявляется в очень лёгкой форме и лишь
в условиях кислородной недостаточности.
Этим лицам противопоказано проживание
в высокогорных районах, служба в авиации,
горноспасательная служба. Вместе с тем
у них наблюдается высокая устойчивость
к малярии (в 13 раз болеют реже, чем
нормальные люди). Невосприимчивость к
малярии у гетерозиготных носителей
гена серповидноклеточной анемии
объясняется неспособностью малярийного
плазмодия осуществить свой жизненный
цикл в эритроцитах с дефектным
гемоглобином. Кодоминирование обычно
выявляется на электрофореграммах
ферментов (рис. 63).
ллельные
гены могут взаимодействовать по типусовместного доминирования
или кодоминирования
(рис. 62). Кодоминирование
- это тип взаимодействия аллельных
генов, при котором оба аллеля проявляют
активность, внося равноценный вклад в
формирование феноти-
па.
Известно, например, что серповидноклеточная
анемия проявляется как аутомно-рецессивное
заболевание в гомозиготном организме.
В таком организме присутствуют два
патологических аллеля одного и того же
гена, контролирующие синтез дефектного
гемоглобина. В гетерозиготном организме
присутствуют нормальный и дефектный
аллельные гены. Причём ни один из них
не доминирует над другим, поэтому в
организме одновременно синтезируются
оба вида гемоглобина (нормальный и
дефектный). У таких индивидов симптоматика
заболевания почти отсутствует или она
проявляется в очень лёгкой форме и лишь
в условиях кислородной недостаточности.
Этим лицам противопоказано проживание
в высокогорных районах, служба в авиации,
горноспасательная служба. Вместе с тем
у них наблюдается высокая устойчивость
к малярии (в 13 раз болеют реже, чем
нормальные люди). Невосприимчивость к
малярии у гетерозиготных носителей
гена серповидноклеточной анемии
объясняется неспособностью малярийного
плазмодия осуществить свой жизненный
цикл в эритроцитах с дефектным
гемоглобином. Кодоминирование обычно
выявляется на электрофореграммах
ферментов (рис. 63).
Кодоминирование проявляется при наследовании групп крови у человека. Особи, имеющие вторую (АА) и третью (ВВ) группы крови, гомозиготны, однако у их гетерозиготных потомков из-за одинакового проявления (экспрессии) генов будет отмечаться четвёртая (АВ) группа крови.
Особым типом взаимодействия аллельных генов является сверхдоминирование, при котором у гетерозигот признак проявляется сильнее, чем у гомозигот. Следовательно, сверхдоминирование - это тип взаимодействия аллельных генов, при котором рецессивный аллельный ген усиливает в гетерозиготном состоянии действие доминантного аллельного гена. Так, при скрещивании чистых линий двух сортов кукурузы гетерозиготное потомство оказывалось более выносливым и продуктивным, чем исходные гомозиготные родительские формы. Однако при дальнейшем самоопылении по мере перехода кукурузы в гомозиготное состояние эти положительные качества гибридов утрачивались. Явление «гибридной мощности» или превосходства гибридов по ряду признаков и свойств над родительскими формами названо Дж. Шеллом в 1914 году «гетерозисом». Сверхдоминирование описано также у дрозофилы, у которой обнаружен рецессивный мутантный ген, гетерозиготы по которому более жизнеспособны, чем гомозиготные мухи диких популяций.