

5. Посттранскрипционные процессы. Процессинг (созревание рнк).
Это совокупность процессов обеспечивающих превращение синтезированной РНК (РНК-транскрипта) в функционально активные РНК (зрелые РНК), которые могут быть использованы при синтезе белков. Сами РНК-транскрипты функционально не активные. Процесс характерен для эукариот.
В результате процессинга изменяется структура и химическая организация РНК. РНК-транскрипт до образования зрелой РНК носит название про-иРНК(или в зависимости от вида РНК – про-тРНК, про-рРНК), т.е. предшественница РНК. Практически все РНК-транскрипты эукариот и прокариот(за исключением иРНК прокариот)подвергаются процессингу. Превращение РНК-транскрипта в зрелую РНК начинается в ядре, когда синтез РНК ещё не закончен и она не отделилась от ДНК. В зависимости от механизмов различают несколько этапов созревания РНК.
Взаимодействие про-иРНК с белком.
Метилирование про-иРНК.
Кэпирование 5’-конца.
Полиаденилирование.
Сплайсинг .
Графическая последовательность этапов изображена на рисунке 58. Следует отметить, что в живых организмах все вышеперечисленные процессы идут параллельно друг другу.
а. Взаимодействие про-иРНК с белком.
У бактерий ещё до окончания транскрипции 5’конец транскрипта сразу же соединяется с рибосомой и иРНК включается в трансляцию. Поэтому, для бактериальной иРНК практически никакая модификация не требуется. У эукариот, синтезированный транскрипт выходит из ядра, попадает в цитоплазму и там соединиться с рибосомой. На своём пути он должен быть ограждён от случайных встреч с сильными реагентами и, в тоже время быть, доступен ферментам процессинга. Поэтому РНК-транскрипт сразу же по мере удлинения взаимодействует с белком. Здесь уместна аналогия – РНК-транскрипт располагается на белке как на операционном столе, он фиксируется химическими связями, одновременно в нём становятся доступными места модификации. РНК, связанная с белком, носит название рибонуклеопротеид ( информосома). В такой форме транскрипт находится в ядре. При выходе из ядра одни РНК продолжают оставаться в соединении с белком, другие выходят из комплекса и принимают участие в трансляции.
б. Метилирование про-иРНК.
Чаще всего происходит у бактерий, у которых имеется специальный аппарат защиты от чужеродной
ДНК (вирусной, фаговой). Этот аппарат состоит из целого ряда ферментов разрезающих чужеродную ДНК или РНК в определённых сайтах в которой находится специфическая последовательность нуклеотидов. Ферменты носят название – рестриктазы. Понятно, что собственный, только что синтезированный РНК-транскрипт, также может быть подвергнут атаке рестриктаз. Чтобы это не случилось специальные ферменты, называемые метилазы,метилируют собственный РНК-транскрипт в тех сайтах, которые могут быть разрезаны собственными ферментами. У эукариот РНК-транскрипт метилируется в меньшей степени.


 Промотор
Терминатор
Промотор
Терминатор
Транскрипция
 Про-иРНК
Про-иРНК


П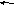 ро-иРНК
фикси-
Белок
ро-иРНК
фикси-
Белок
 рванная
на белке
рванная
на белке
М етилирование
про-иРНК
етилирование
про-иРНК

К эпирование
про-иРНК
эпирование
про-иРНК


П







 олиаденилирование
олиаденилирование

Сплайсинг (см. далее)
Рис. 58. Схема основных моментов процессинга.
в. Кэпирование 5’конца.
Заключается в химическом и конформационном изменении
5’конца синтезированной РНК. Кэпирование происходит в момент синтеза РНК, ещё до её отделения. Процесс заключается в присоединении к свободному концу про-РНК специальных химических веществ, которые изменяют конформацию концевого участка. Кэпирование необходимо для инициации процесса трансляции.
Специальные ферменты присоединяют к 5’концу про-иРНК ГДФ (гуанозиндифосфат), а затем метилируют его.
ГДФ


 5’
про-иРНК
5’
про-иРНК

СН3

КЭП = ГДФ + СН3
Рис.59. Структура КЭПа на 5’конце пре-иРНК эукариот.
Функции КЭПа.
Инициирует синтез белка.
Предохраняет про-иРНК от распада.
Участвует в удалении интронов.
г. Полиаденилирование.
Это процесс присоединения к 3’ концу про-иРНК 100 – 200 остатков адениловой кислоты. Эти остатки носят название поли-А последовательности (поли-А хвосты). Полиаденилированию подвергаются не все про-иРНК. Например, молекулы всех типов гистонов не содержат поли-А последовательности. Полиаденилирование предохраняет иРНК от разрушения.
На растущей цепи и-РНК имеется специальная последовательность нуклеотидов (ААУААА). Особый фермент ( полиА-полимераза) находит это сочетание нуклеотидов, разрезает про-иРНК в этом месте и формирует полиадениловый хвостик.
Значение поли –А последовательностей:
Облегчают выход иРНК из ядра в цитоплазму.
Предохраняют иРНК от разрушения.
Недавно было выявлено ещё одно интересное свойство поли-А последовательностей – они участвуют в терминации синтеза про-иРНК. РНК-полимераза, формируя последовательность ААУААА в про-иРНК, получает сигнал о завершении синтеза РНК-транскрипта. Но синтез сразу не прекращается. Полная остановка его наступает после того, как РНК-полимераза встречает на матричной нити ДНК специфическую последовательность нуклеотидов (у разных генов она разная), которая и даёт окончательный сигнал о прекращении синтеза РНК.
ГТФ ПолиА - последовательность



 рАрАрАрАрАрАрАрА-ОН
рАрАрАрАрАрАрАрА-ОН

 СН3
СН3
КЭП = ГТФ + СН3
Рис. 60. Структура КЭПа на 5’конце про-иРНК эукариот и полиадениловая последовательность на 3’конце про -иРНК.
д. Сплайсинг.
В РНК-транскрипте содержится определённое количество нуклеотидных последовательностей, которые были необходимы для успешного завершения трансляции и последующей модификации транскрипта (кэпирования, полиаденилирования и т.д.). Для выполнения основной роли РНК в цитоплазме – трансляции, эти последовательности не только не будут иметь функционального значения, но могут помешать нормальному течению синтеза белка. Поэтому в клетке предусмотрен механизм освобождения первичного транскрипта от целого ряда последовательностей, не имеющих решающего значения в трансляции.
К таким последовательностям прежде всего относят интроны.
Ген, с которого транскрибировалась про-иРНК содержит кодирующие и некодирующие последовательности. Кодирующие последовательности гена определяют аминокислоту и их последовательность в белке. Не кодирующие последовательности таким свойством не обладают. Кодирующие и некодирующие последовательности в гене чередуются, и их количество зависит от индивидуальных генов. В первичном транскрипте также содержатся кодирующие и некодирующие последовательности. Такая организация генов и про-РНК характерна для эукариот. Некодирующие последовательности про-иРНК носят название интроны, а кодирующие –экзоны. Длина интронов может быть от 50 до 12000 нуклеотидов. Ген начинается и
кончается экзоном. Прерывистое строение гена характерно для большинства эукариот. Интроны могут содержать все виды РНК – иРНК, тРНК, рРНК.
Вся совокупность экзонов (кодирующих белки) в геноме человека занимают всего 1,1 – 1,4 %. Средний ген человека содержит 9 интронов. По мере упрощения
организации организмов совокупная величина их экзонов возрастает (например у бактерий она равна 86%).
В вырезании интронов из РНК-транскрипта и сшивании оставшихся экзонов, принимает участие многокомпонентный комплекс. Основными его составляющими являются малые ядерные РНК (мяРНК) и белки-ферменты.
В целом комплекс носит название малые ядерные рибонуклеопротеиды, мяРНП или сплайосома. Сам процесс достаточно сложен и состоит из нескольких этапов (см. рис. 58).
1. Формирование сплайосомы. К началу и концу интрона прикрепляются фрагменты белка и мяРНК (рис. 56, Д) формируя сплайосому. (рис. 56, Д) Прикрепление комплекса мяРНП (рис. 56, Е).
 Экзон 1 Интрон
Экзон 2
Экзон 1 Интрон
Экзон 2
А 



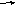

 Д
Д
Б

Е
В
Петля
 интрона вырезана
интрона вырезана
Г
 Ж
Ж
Рис. 61. Схема сплайсинга (объяснение в тексте).
Сближение соседних экзонов, за счёт образования петли интрона. Разрезание на границе экзон-интрон и соединение соседних (первого и второго) экзонов(рис. 56, В).
Удаление и разрушение петли и сплайосоме (рис. 56, Г, Ж).
Необходимо отметить, что при повреждении (мутации) интрона сплайсинг может быть не закончен, интрон не вырезан и конченый продукт – иРНК будет нести несвойственные ей последовательности нуклеотидов. Понятно, что это может привести к нарушению трансляции и выключению из метаболизма определённого белка
е. Альтернативный сплайсинг.
Такой тип сплайсинга происходит при экспрессии одного и того же гена в разных тканях.
Сущность его в том, что один и тот же участок гена в разных тканях может выступать в качестве интрона и экзона. Это приводит к образованию разных иРНК, которые кодируют белки с различной ферментативной активностью.
Так в клетках щитовидной железы синтезируется гормон кальцитонин. Он тормозит высвобождение кальция из костей. Ген, контролирующий синтез каль-
Ген, контролирующий кальцитонин

э и э и э и э и э и э
Д





 НК
НК
1 2 3 4 5 6
э и э и э и э и э и э







про-иРНК
1 2 3 4 5 6



 В щитовидной железе
В клетках головного мозга
В щитовидной железе
В клетках головного мозга
























иРНК
 1 2 3 4 1 2
3 5 6
1 2 3 4 1 2
3 5 6



Кальцитонин Кальцитонинподобный белок
Рис.62. Альтернативный сплайсинг кальцитонина и кальцитонин-подобного белка.
цитонина, состоит из 6 экзонов, первичный транскрипт этого гена ( про-иРНК) также состоит из 6 экзонов (рис. 62). Из первичного транскрипта формируется зрелая иРНК содержащая 4 экзона – 1,2,3,4. Экзоны № 5 и 6 были прочитаны как интроны и вырезаны. На основе такой и РНК синтезируется кальцитонин. В клетках головного мозга из первичного транскрипта, содержащего 6 экзонов, формируется зрелая иРНК, состоящая из 5 экзонов – 1,2,3,5,6. Четвёртый экзон был вырезан как интрон. Такая иРНК контролирует синтез кальцитонинподобного белка, отвечающего за вкусовое восприятие.
Другой ген Icarus ( в названного в честь легендарного Икара) способен обеспечить за счёт альтернативного сплайсинга синтез 6 различных полипептидов. Кроме этого полипептиды образуют между собой в клетке около 20 различные ансамбли из одних и тех же полипептидов или различных.
Нарушение механизма сплайсинга может привести к патологическим состояниям, которые носят общее название талассемии. К ним относят заболевания связанные с частичным или полным подавление синтеза одной из цепей гемоглобин ( α- или β-цепей). Например, болезни, связанные с недостатком синтеза β -цепи гемоглобина, могут возникнуть в результате мутаций в двух участка гена, кодирующего β-цепь – в сайте ответственном за полиаденилирование и в одном из интронов. В первом случае нарушается процесс формирования полиаденилового хвостика и формируется неполноценная β-цепь гемоглобина. Во втором случае сплайосома не способна вырезать повреждённый интрон и зрелая иРНК β-цепи гемоглобина не образуется. В любом случае нормальная функция эритроцитов будет существенно нарушена.
МЗ. Процессинг (или созревание РНК) это процесс превращения только что синтезированной, не активной РНК (про-иРНК) в функционально активную РНК. Процесс связан со структурными и химическими модификациями про-иРНК. Происходит в ядре до момента выхода РНК в цитоплазму. Состоит из нескольких этапов: присоединение про-иРНК к белку, метилирование некоторых оснований, маркировка одного из концов, полиаденилирование другого (противоположного) конца, вырезания интронов и сшивание экзонов. Последние два процесса носят название сплайсинг.
Вопросы к экзаменам.
1. Каким образом ферменты определяют большинство мест, где имеется повреждение молекулы ДНК?
ОТВЕТ. В месте повреждения молекулы ДНК в большинстве случаев наступает локальная денатурация. Её и определяют ферменты.
2. Что происходит в месте повреждения молекулы ДНК ?
ОТВЕТ. В месте повреждения наступает локальная денатурация.
3. На основании чего ферменты репарации восстанавливают необходимую последовательность нуклеотидов в месте повреждения одной нити ДНК?
ОТВЕТ. На основании принципа комплементарности к нуклеотидам оппозитного участка нити ДНК.
4. На основании чего ДНК-полимераза правильно застраивает нуклеотидами бреши в повреждённой нити ДНК ?
ОТВЕТ. На основании принципа комплементарности нуклеотидов застраиваемой цепи к нуклеотидам оппозитной нити.
5. Какой тип репарации осуществляется ферментом, который активируется фотоном?
ОТВЕТ. Фотореактивация.
6. Какой фермент осуществляет репарацию используя энергию солнца?
ОТВЕТ. Фотолиаза.
Какой фермент принимает непосредственное участие в синтезе молекуле РНК?
ОТВЕТ. ДНК-зависимая РНК-полимераза или РНК-полимераза.
Перечислите периоды транскрипции.
ОТВЕТ. Инициация, элонгация, терминация.
Из каких компонентов состоит инициаторный комплекс в процессе транскрипции?
ОТВЕТ. Из специального белка осевшего на промотор, РНК-полимеразы и транскрипционных факторов.
9. Как называется участок ДНК, где формируется инициаторный комплекс в процессе транскрипции?
ОТВЕТ. На промоторе.
10. Как называется последовательность нуклеотидов у прокариот, которую определяет специальный белок осаждающий на промоторе в период инициации транскрипции?
ОТВЕТ. Блок Прибнова.
11. Как называется последовательность нуклеотидов у эукариот, которую определяет специальный белок осаждающий на промоторе в период инициации транскрипции?
ОТВЕТ. ТАТА-бокс.
12. Где в молекуле ДНК располагается блок Прибнова у прокариот?
ОТВЕТ. На промоторе.
13. Где в молекуле ДНК располагается ТАТА-бокс у эукариот?
ОТВЕТ. На промоторе.
14. Как называется ферментативный комплекс, который формирует транскрипционный глазок?
ОТВЕТ. Инициаторный комплекс.
15. Как называется участок молекулы ДНК с которого начинается синтез РНК ?
ОТВЕТ. Стартовой точкой, сайт начала транскрипции.
16. Назовите нуклеотиды, которые находятся в терминаторе и возможно участвуют в прекращении транскрипции.
ОТВЕТ. Г,Ц.
17. Назовите вторичную структуру в терминаторе, которая возможно участвует в прекращении транскрипции,
ОТВЕТ. Шпилька.
18. Как называются кодоны находящиеся в терминаторе и возможно участвующие в прекращении транскрипции.
ОТВЕТ. Бессмысленные (нонсенс) кодоны.
Без медицины, НУЖНО ещё процессинг и далее но я не успел это прочитать.