

беспорядочно.
Хорошо известно также, что нервные волокна проходят часто длинный путь по различным тканям от нервного центра к рецептору или эффектору и «узнают» их. Скорее всего нервные окончания растут по микроструктурам субстрата, как они делают это и в культуре тканей. Субстратом для движения может быть и соседняя клетка, если оболочка ее натянута. А так как поверхность направленно движущейся клетки сама вытягивается, то и она может служить субстратом для движения и поляризации следующей за ней клетки.
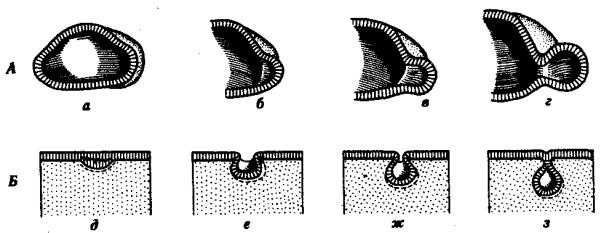
Рис. 8.2. Перемещение клеточных пластов.
А —путем выпячивания на примере образования глазного пузырька; Б—путем впячивания на примере образования слухового пузырька:
а — стенка переднего мозга, б —местное ускорение роста, в — выпячивание, г — глазной пузырек, д—ж —углубление ямки, з —отшнуровывание пузырька
Согласованные перемещения пластов эпителиальных клеток также изучаются. К ним относятся изгибы клеточных пластов путем выпячивания или впячивания, отшнуровка, образование утолщений — плакод (рис. 8.2).
Таким, образом, несомненно, что для миграции клеток очень важны их способность к амебоидному движению и свойства клеточных мембран. И то, и другое генетически детерминировано, так что и сама миграция клеток находится под генетическим контролем, с одной стороны, и влияниями окружающих клеток и тканей — с другой.
8.2.3.Сортировка клеток
Впроцессе эмбриогенеза клетки не только активно перемешаются, но и «узнают» друг друга, т.е. образуют скопления и пласты только с определенными
клетками. Значительные координированные перемещения клеток характерны для периода гаструляции. Смысл этих перемещений заключается в образовании обособленных друг от друга зародышевых листков с совершенно определенным
363

взаимным расположением. Клетки как бы сортируются в зависимости от свойств, т.е. избирательно.
Замечено, что необходимым условием сортировки являются степень подвижности клеток иособенности их мембран. В поздней бластуле амфибий, например, клетки будущей эктодермы обладают тенденцией слипаться друг с другом и распространяться в виде сплошного слоя над мезодермой и энтодермой. Эта тенденция проявляется и в культуре тканей. Клетки мезодермы имеют тенденцию впячиваться в любой находящийся поблизости комок клеток, а клетки энтодермы относительно неподвижны.
Эти особенности трудно объяснить, если наблюдать только за нормальным течением развития. Впервые на различия в подвижности и способности к слипанию (адгезии) между клетками, относящимися к разным зародышевым листкам, обратил внимание Г. Гольтфретер в 30-х гг. XX столетия. Затем производили модификации его опытов с самыми разными клетками. Смысл этих опытов состоит в том, что зародыши тритонов или других животных на стадии гаструлы диссо-циируют с помощью фермента трипсина, который разрушает материал, соединяющий клетки друг с другом. Диссоциированные (дезагрегированные) клетки тщательно перемешивают и затем создают такие условия, чтобы клетки могли свободно перемещаться и воссоединяться друг с другом. Сначала клетки представляют собой беспорядочную смесь, затем клетки эктодермы, мезодермы и энтодермы разделяются (сегрегируют) и собираются в отдельные группы, каждая из которых представляет собой клеточный агрегат из однородных клеток. Образуются снова зародышевые листки, располагающиеся иногда даже в обычном для них порядке (рис. 8.3).
Рис. 8.3. Адгезия клеток зародышевых листков:
а—смесь диссоциированных клеток гаструлы амфибий, б—клетки эктодермы, мезодермы и энтодермы, группирующиеся послойно путем адгезии
Подобную агрегацию клеток зародышевого листка с себе подобными можно объяснить способностью к избирательному слипанию клеток одного типа между собой. Одновременно это является проявлением ранней дифференцировки клеток на стадии гаструлы.
Существует ряд гипотез, объясняющих избирательную сортировку клеток. Возможно, что контакты между подобными клетками сильнее, чем между чужеродными клетками, из-за различий в поверхностном заряде их мембран. Обнаружено, что поверхностный заряд клеток мезодермы ниже, чем клеток экюили энтодермы, благодаря чему клетки мезодермы легче деформируются и втягиваются в бластопор в начале гаструляции. По другой гипотезе, контактные
364
взаимодействия между одинаковыми клетками основываются на антигенных свойствах их мембран.
Избирательная адгезия клеток определенного зародышевого листка друг с другом является необходимым условием нормального развития. Это показано путем сопоставления двух процессов: течения гаструляции в естественных условиях и способности клеток определенного зародышевого листка агрегировать в комплекс с себе подобными. Показано, что если у межвидовых гибридов зародышей амфибий нарушена гаструляция, то диссоциированные в опыте клетки зародышевых листков таких гибридов не способны к агрегации с себе подобными, если же в некоторых случаях гибридам удавалось образовать гаструлу, то и в эксперименте диссоциированные клетки зародышевых листков были способны к реагрегации. Повидимому, в обеспечении сортировки клеток важное место принадлежит генетическим механизмам.
Одним из примером потери клетками способности к избирательной сортировке и слипанию является их беспорядочное поведение в злокачественной опухоли.
Таким образом, сортировка клеток и их избирательная адгезия наряду с другими клеточными процессами играет важную роль в морфогенезе развивающегося зародыша и одновременно подвержена многоуровневым регуляционным воздействиям (генетическим, межклеточным, онтогенетическим), отражая целостность организма как системы.
8.2.4.Гибель клеток
Вразвитии зародышей наряду с размножением клеток важную роль играют
процессы гибели клеток.
Внастоящее время различают два принципиально различных типа клеточной гибели: апоптоз (в переводе с греческого «отпадающий») и некроз.
Апоптоз широко распространен и типичен для физиологических условий. Наряду с описанными выше делением, сортировкой и миграцией клеток, он способствует достижению характерных для определенного биологического вида черт его морфофункциональной организации. Следовательно, апоптоз является естественным, эволюционно обусловленным и генетически контролируемым механизмом морфогенеза. Некроз клеток возникает в нефизиологических условиях, например, в связи с действием неблагоприятных факторов, таких, как стойкое кислородное голодание, разного рода токсины и другие. Некроз обычно сопровождается воспалением и является патологическим процессом. Указанные воззрения в отношении апоптоза и некроза сформировались за последние три десятилетия и поэтому еще нередко некоторые физиологические процессы клеточной гибели описываются термином «некроз» вместо термина «апоптоз».
Наиболее яркие примеры разрушения клеток и органов относятся к постэмбриональным стадиям метаморфоза земноводных и насекомых. У головастиков резорбируются (рассасываются) хвост, кишечник и жаберные крышки,
365
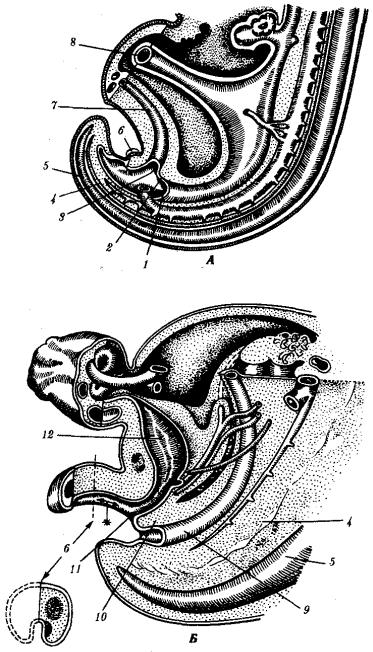
у личинок насекомых разрушается большинство внутренних органов. В ходе эмбрионального развития высших позвоночных и человека также имеют место процессы дегенерации органов, которые вначале закладываются, а затем исчезают. У особей женского пола дегенерируют вольфовы протоки, у особей мужского пола —мюллеровы протоки, что является, по-видимому, результатом влияния половых гормонов. У эмбриона человека вначале закладываются ребра у 7-го шейного позвонка и 9—10 хвостовых позвонков, затем они обычно исчезают, так что шейные позвонки, как правило, ребер не несут и в копчике остается 4—5 позвонков (рис. 8.4).
Рис. 8.4. Область клоаки и хвоста у зародышей человека.
366
А —в конце 4-й недели; Б—на 8-й неделе: 1—ректальная область клоаки, 2— уроректальная складка, 3—область мочеполового синуса в клоаке, 4—хорда, 5— нервная трубка, 6—половой бугорок, 7—аллантоис, 8—желточный стебелек, 9— прямая кишка, 10—уроректальная перегородка, 11—мочеполовой синус, 12— мочевой пузырь
Немалое значение принадлежит процессам гибели клеток при образовании полостей тела или сосудов (так называемая кавитация), имеющих вначале вид тяжей без просвета.
В центральной нервной системе сначала образуется больше нервных клеток, чем затем сохраняется, так как часть нейронов, не установивших связи со своими мишенями, погибает.
Наиболее изучены процессы гибели клеток при образовании дефинитивной формы конечностей птиц и млекопитающих. У цыпленка почки конечностей закладываются в виде боковых утолщений соматоплевры примерно на 55-м часу развития. Они вырастают из тела зародыша в виде выступов, покрытых эктодермой
изаполненных мезодермальной тканью. По мере их роста начинают проступать контуры конечностей. Процесс формирования контуров сопровождается отмиранием клеток в ряде мезодермальных участков почки конечности. Участки будущей гибели клеток выявлены с помощью прижизненных красителей.
Так называемая задняя некротическая зона (ЗНЗ) обеспечивает формирование контуров проксимальных областей конечностей. Максимальной протяженности ЗНЗ достигает к 96-му часу развития. К этому времени гибнет 1500—2000 клеток, поглощаемых примерно 150 макрофагами. Другие клетки мезодермы, примыкающие к зоне некроза, не погибают. Группа клеток будущей ЗНЗ, пересаженная со своего местонахождения на боковую поверхность тела зародыша за 40 ч до начала ожидаемого некроза, все равно погибает в те же сроки, что и на своем обычном месте. В данном примере проявляется генетическая запрограммированность гибели определенных клеток, которую можно образно назвать «внутренними часами смерти».
Если клетки ЗНЗ пересадить на дорсальную поверхность почки крыла, то ее клетки не погибнут. Следовательно, некоторые другие клетки способны предотвратить гибель клеток ЗНЗ. Такой способностью обладают клетки мезодермы из дорсальной ненекротической области крыла. Показано, что контакт клеток ЗНЗ с «защищающими» их от гибели клетками дорсальной мезодермы крыла должен продолжаться не менее 6 сут., после чего они могут уже продолжать существование
ибез контакта с этими клетками. Таким образом, гибель клеток контролируется не только генетически, но и на уровне клеточных взаимодействий. Очевидно, что в описанных событиях мы наблюдаем апоптоз, несмотря на сохранение в названиях гибнущих зонах — «некротические».
Интересно сравнение влияния условий на процессы гибели клеток в дистальной части лапок у цыпленка и утенка. Как известно, у утят гибель клеток в дистальной части лапки невелика, вследствие чего у них между вторым, третьим и четвертым пальцами образуются перепонки. Когда создавали химеры, соединяя
367
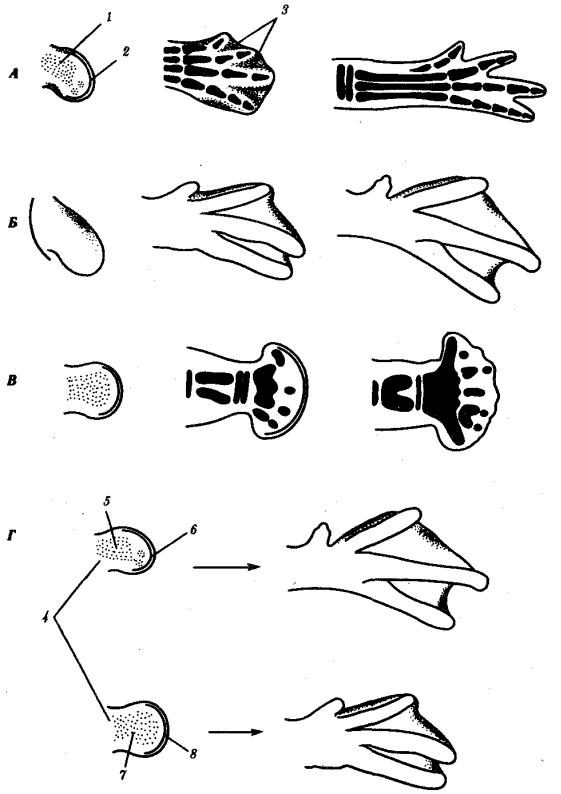
мезодерму из почки ноги утки с эктодермой цыпленка и наоборот, подсаживая затем химерные почки конечностей на боковую поверхность куриного зародыша, в обоих случаях получали развитие по типу утиной конечности, т.е. с перепонкой между пальцами (рис. 8.5).
368