

ТОХФ / 6 группа (ХТБМП) / Биохимия / Лекции - Коваленко - 2003 / 12. анаболизм
.doc
ОСНОВНЫЕ АНАБОЛИЧЕСКИЕ ПРОЦЕССЫ
1. Глюконеогенез
Главным быстро мобилизуемым источником энергии в клетках является глюкоза, запа-сы которой находятся в виде продуктов ее поликонденсации – крахмала, гликогена или других полисахаридов. В крови человека поддерживается постоянный уровень глю-козы около 5 ммоль/л, но небольшие отклонения от этого нормального уровня возмож-ны при интенсивной работе или после еды. Диффузия глюкозы в клетки идет по градиенту концентраций и есть специальный мембранный белок, облегчающий этот процесс. Уровень глюкозы в крови поддерживается с помощью гормона поджелудоч-ной железы инсулина и ряда других гормонов. Инсулин имеет пептидную природу, он состоит из 51 аминокислоты, но секреторные клетки (островки Лангерганса) произ-водят проинсулин, который у разных животных содержит от 78 до 86 аминокислотных компонент в виде одной цепочки с тремя дисульфидными связями. Активация с обра-зованием самого инсулина происходит за счет гидролитического расщепления молеку-лы проинсулина с элиминированием из нее центрального участка (С‑цепь) с сохране-нием всех дисульфидных связей в активном двухцепочечном полипептиде (А- и В‑цепи). Инсулин не проходит через клеточную мембрану, но он связывается с белком переносчиком глюкозы и ускоряет ее диффузию в клетку. При избытке глюкозы она превращается в гликоген, а при недостатке глюкозы она образуется из гликогена. При недостатке инсулина или при нарушении транспорта глюкозы в клетки организм повы-шает ее концентрацию в крови для того, чтобы увеличить движущую силу диффузии за счет градиента концентраций. Но это приводит и к более интенсивному выделению глюкозы почками (сахарный диабет). Испытывающие дефицит по глюкозе клетки начинают синтезировать ее из аминокислот сократительных белков, подключая к это-му окислительные превращения жирных кислот. В результате этого мышечные ткани атрофируются и разрастается жировая ткань.
Если АТФ является универсальным источником энергии, то глюкоза представляет собой главный источник быстро мобилизуемой энергии. Для нервной системы, почек, семенников, тканей эмбриона это к тому же единственный источник АТФ. Глюкоза –это единственный источник энергии, который легко преодолевает гематоэнцефаличес-кий барьер даже у диабетиков; человеческий мозг потребляет около 120 г глюкозы в cутки.

Роль глюкозы очевидна, но возможности запасания ее в виде гликогена ограничены и в организме взрослого человека содержится всего лишь несколько сот граммов этого полисахарида. Поступление глюкозы с пищей (в основном в виде крахмала) не всегда соответствует действительным потребностям и поэтому организм животных в значи-тельной мере зависит от образования глюкозы из поступающих с пищей веществ. Не-которые впадающие в спячку животные к началу зимы запасают столько жира, что он обеспечивает обогрев и обменные процессы в течение нескольких холодных месяцев, а перелетные птицы запасают в жировой ткани энергию, необходимую для перелета на многие тысячи километров. Однако в организмах подавляющего большинства тепло-кровных избыток пищевых веществ закладывается в жировую ткань, а при недостатке глюкозы в пище на ее образование расходуются белки.
Напомним три основных правила протекания метаболических процессов:
-
Катаболические и анаболические пути одного вещества различны. Это значит, что есть крайней мере одна стадия, по которой они не совпадают.
-
Катаболический и анаболический путь превращения одного вещества контролиру-ется различными регуляторными ферментами, но регуляция биосинтеза и биодегра-дации одного вещества взаимосвязаны. Активация биосинтеза приводит к подавле-нию катаболического превращения.
-
Анаболические процессы расходуют АТФ, который поставляется катаболическими. Поскольку КПД всегда менее 100 %, затраты АТФ оказываются выше, чем это сле-дует из термодинамических расчетов. Это обеспечивает необратимость анаболичес-ких превращений.
При изучении процесса гликолиза было установлено, что конечный продукт анаэроб-ного превращения глюкозы – молочная кислота – снова превращается в глюкозу. При этом около 1/5 молочной кислоты окисляется до диоксида углерода.
В процессе гликолиза глюкозы образованию молочной кислоты предшествует катали-зируемое пируваткиназой образование пирувата из фосфоенолпирувата с генериро-ванием АТФ:

По энергетическим соображениям обращение этого пути невозможно, так как прямая реакция сопровождается выделением 7,5 ккал/моль. Поэтому для глюконеогенеза обра-зование фосфоенолпирувата из пирувата включает обходной путь. Сначала пируват биотинзависимым ферментом карбоксилируется с образованием оксалоацетата. Этот процесс идет в митохондриях и для переноса оксалоацетата в цитозоль, где идет биосинтез глюкозы, оксалоацетат восстанавливается в малат, для которого существует специальная транспортная система, и уже в цитозоле малат дегидрируется с образова-нием оксалоацетата. Анион щавелевоуксусной кислоты фосфорилируется действием ГТФ в присутствии фосфоенолпируваткарбоксикиназы с одновременным декарбокси-лированием:

Хотя изменение стандартной свободной энергии при этом равно + 0,2 ккал/моль, в реальных условиях клетки этот процесс сопровождается выделением 6 ккал/моль, что делает его необратимым.
В обращение процессов гликолиза фосфоенолпируват гидратируется и восстанавлива-ется до фосфоглицеринового альдегида, половина которого превращается в фосфат диоксиацетона, после чего под действием альдолазы они превращаются в 1,6-дифосфат фруктозы. Если гликолиз инициируется фосфорилированием глюкозы и фосфофрук-тозы аденозинтрифосфатом, то обратный процесс образования АТФ из дифосфата фруктозы или фосфоглюкозы и АДФ по термодинамическим соображениям невозмо-жен и фосфатные группы отщепляются гидролитически.
Если суммарный процесс гликолиза представляется схемой
![]() ,
,
то на обратный процесс затрачивается уже втрое больше АТФ, чем образуется в прямом:
![]()
Пируваткарбоксилаза это аллостерический фермент, активируемый ацетилкофермен-том А, то есть при накоплении в клетке неиспользуемого ацетилкофермента А начина-ется усиленный биосинтез глюкозы из пирувата.
В глюкозу и далее в гликоген могут превращаться все промежуточные продукты цикла Кребса. Большинство аминокислот также относятся к глюкогенным. В их числе ала-нин, глютаминовая, аспарагиновая кислота и их амиды, превращающиеся (амиды после гидролиза) трансаминазами в пируват, кетоглутарат и оксалоацетат, кроме того, эти кетокислоты образуются и из других аминокислот. Особенно интенсивно идет превращение аминокислот в глюкозу при недостатке инсулина.
В нормальных условиях ацетилкофермент А не может превращаться в пируват и по-этому жирные кислоты с четным числом атомов углерода в глюкозу не превращаются.
-
Биосинтез жирных кислот
Как биодеградация (-окисление), так и биосинтез жирных кислот протекают с участи-ем двухуглеродных фрагментов ацетилкофермента А. На начальном этапе протекает катализируемая ацетил-СоА-карбоксилазой реакция карбоксилирования ацетилкофер-мента А с образованием малонилкофермента А:
![]()
Биотинзависимый фермент ацетил-СоА-карбоксилаза – это регуляторный фермент, определяющий скорость образования высших кислот. Положительным эффектором ацетил-СоА-карбоксилазы служит цитрат. Смысл этой предварительной реакции заключается в том, чтобы повысить СН-кислотность метаболита и облегчить реакцию наращивания углеродной цепи в следующей далее реакции С-ацилирования, протекаю-щей по типу реакции сложноэфирной конденсации.
Биосинтез жирных кислот осуществляется в мультиферментной системе, состоящей из семи ферментов. Растущая ацильная цепь связана ковалентной связью с ацилпере-носящим белком (АПБ), занимающим центральное положение в этой системе. Как и в коферменте А реакционный центр АПБ представлен сульфгидрильной группой цисте-амина, который ацилирован по аминной группе пантотеновой кислотой, но в отличие от кофермента А этот фрагмент АПБ (АПБ-SH) связан фосфатной группой не с адено-зиновой составляющей, а с гидроксильной группой серинового фрагмента в молекуле АПБ:

В синтезе жирных кислот принимает участие еще одна сульфгидрильная группа молекулы АПБ (Cys-SH), но она принадлежит цистеиновому фрагменту белковой цепи. Тогда этот фермент (энзим, Е) вместе с его простетическими группами может быть схематически изображен следующим образом:

Синтез жирной кислоты на мультиферментном комплексе начинается с переноса ацетильной группы с ацетилкофермента А на цистеиновую тиольную группу синтазы жирных кислот, который катализируется АПБ-ацетилтрансферазой:

Затем АПБ-малонилтрансфераза переносит на вторую сульфгидрильную группу оста-ток малоновой кислоты с малонил-СоА:

Теперь ацетильная группа по схеме внутримолекулярной сложноэфирной конденсации переносится с цистеинового остатка на фрагмент малоновой кислоты с декарбокси-лированием. Этот процесс катализируется 3-кетоацил-АПБ-синтазой:

То есть трехуглеродный фрагмент малоновой кислоты не полностью участвует в био-синтезе жирных кислот. Он теряет карбоксилатный фрагмент, который снова вклю-чается в процесс для активации метильной группы другой молекулы СоА-S-Ac.
В последующих превращениях происходит восстановление ацетоацетильного фрагмен-та. Сначала NADPH восстанавливает кетогруппу до гидроксильной группы с образова-нием D-3-гидроксибутирилзамещенного АПБ:

Интересно, что при -окислении жирных кислот гидроксиацилкофермент А образуется в результате гидратации двойной связи и асимметричный атом углерода с гидроксиль-ной группой имеет L-конфигурацию, а при биосинтезе жирных кислот образуется D‑3‑гидроксибутирилзамещенный АПБ, который дегидратируется с образованием транс-2-бутеноил-АПБ. Из этого можно сделать вывод о том, что гидратазы ката-болического и анаболического путей различны, даже с учетом обратимости реакций гидратации или дегидратации, так как они различаются по одному из субстратов. При восстановлении транс-2-бутеноил-АПБ образуется бутирил-АПБ:

После этого бутирильный фрагмент переносится на цистеиновую сульфгидрильную группу и приведенный выше цикл превращений повторяется с ацилированием мало-нильного остатка уже не ацетильным, а бутирильным фрагментом. Этот процесс по-вторяется до тех пор, пока длина ацильного фрагмента не достигнет шестнадцати ато-мов углерода, то есть пока не образуется пальмитиновая кислота. Это конечный про-дукт биосинтеза в этом комплексе ферментов. Все остальные жирные кислоты обра-зуются в результате наращивания или укорочения цепи пальмитиновой кислоты и реакций дегидрирования. Суммарно образование пальмитиновой кислоты выражается уравнением:
![]()
![]()
Строение мультиферментного комплекса, на котором протекают реакции биосинтеза пальмитиновой кислоты, обычно представляется следующей схемой:
-

Е1 - АПБ,
Е2 - ацетилтрансфераза,
Е3 - малонилтрансфераза,
Е4 - 3-кетоацил-АПБ-синтаза,
Е5 - 3-кетоацил-АПБ-редуктаза,
Е6 - 3-гидроксиацил-АПБ-дегидратаза,
Е7 - еноил-АПБ-редуктаза.
однако в действительности шесть ферментов от Е2 до Е7 расположены по полукругу, а этот мультиферментный комплекс состоит из двух одинаковых систем вместе действительно организованных в форме круга.
3. Биосинтез терпеноидов
Еще один анаболический путь превращения ацетил-кофермента А представлен биосин-тезом терпеноидов. Он начинается с образования ацетоацетилкофермента А по схеме сложноэфирной конденсации, катализируемой тиолазой:
![]()
Еще одна молекула ацетил-кофермента А реагирует с ацетоацетилкоферментом А по схеме альдольной конденсации с образованием 3-гидрокси-3-метилглутарилкофермен-та А при катализе соответствующей синтазой:

Теперь гидроксиметилглутарил-СоА-редуктаза переводит этот тиоэфир дикарбоновой кислоты в диоксикарбоновую кислоту, называемую мевалоновой кислотой:

Соответствующие киназы переносят на гидроксильные группы этого вещества один фосфатный и один пирофосфатный фрагмент от трех молекул АТФ:
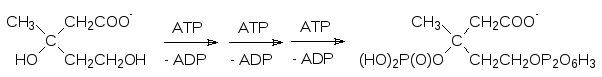
Такая фосфорилированная мевалоновая кислота отщепляет фосфат и декарбоксилиру-ется с образованием изопентенилпирофосфата:
 ,
,
который находится в равновесии с диметилаллилпирофосфатом. Далее идет последо-вательное алкилирование изопентенилпирофосфата диметилаллилпирофосфатом с образованием геранилпирофосфата и далее с образованием фарнезилпирофосфата:


Теперь две молекулы фарнезилпирофосфата соединяются по атомам углерода, несу-щим пирофосфатные остатки (промежуточным продуктом является прескваленпиро-фосфат с циклопропановым фрагментом), и образуется сквален:

Сквален окисляется скваленэпоксидазой с образованием 2,3-эпоксида, который в активном центре скваленоксидциклазы сворачивается в структуру, обеспечивающую образование тетрациклического ланостерина в результате протонирования и раскрытия эпоксидного цикла:

Образовавшийся в результате циклизации карбокатион стабилизируется с отщепле-нием протона с одновременной миграцией двух метильных групп и превращается в ланостерин:

Далее идут окислительные превращения с отщеплением метильных групп, гидрирова-нием двойных связей, дегидрированием с образованием двойных связей и другие ре-акции, приводящие к образованию холестерина, эргостерина и других стероидных липидов, витамина D, стероидных гормонов, желчных кислот.
Изопентенилпирофосфат является также базовым веществом для образования вита-минов А, Е и К, каротиноидов, каучука и других полиизопренов, фитольной компо-ненты хлорофилла и других полипренольных компонент, например в убихиноне.