

ТОХФ / 6 группа (ХТБМП) / Биохимия / Лекции - Коваленко - 2003 / 13. н-г регуляция
.doc
Нейро-гуморальная регуляция
Многоклеточные организмы, состоящие из огромного числа различных типов тканей и органов, нуждаются в регуляции взаимодействия между ними для нормального функ-ционирования в нестационарных условиях. В основе этой регуляции всегда лежат молекулярные механизмы. Превращения субстратов регулируются ферментами, и повышение концентраций субстратов по закону действующих масс приводит к уско-рению их превращений. Кроме этого, регуляторные ферменты изменяют каталитичес-кую активность под действием эффекторов – ингибиторов или активаторов, – а это уже регуляция по принципу обратной связи. Однако только такими простыми взаимосвя-зями нельзя обеспечить даже простое существование сформировавшего многоклеточ-ного организма в постоянно изменяющихся условиях, и уж тем более его развитие с дифференциацией эмбриональных клеток в различные органы и ткани.
Более сложная регуляция, которая также основана на принципах обратной связи, осу-ществляется нервной системой с участием сигнальных веществ, поскольку иннервация каждой клетки организма несколькими типами нервных окончаний просто невозможна. В общем случае эти регуляторные молекулы можно классифицировать по их функциям на три группы:
-
гормоны
-
факторы роста и цитокины
-
нейромедиаторы
Более всего известны классические сигнальные молекулы – гормоны. Многие из них содержатся в организме в сравнительно больших количествах и поэтому они были вы-делены и стали известны уже давно. У животных гормоны образуются в специальных клетках, собранных в железы внутренней секреции (эндокринные железы), которые выделяют их прямо в кровь (у насекомых гемолимфа). Этим они отличаются от других железистых органов, выделяющих свои секреты в протоки, которые так или иначе связаны с внешней средой (поджелудочная железа – экзокринная и эндокринная функ-ция). Разносимые кровью по всему организму гормоны находят свои клетки-мишени на значительном удалении от секретирующей их железы.
Факторы роста – это регуляторные молекулы (у животных это белки). Они отличаются от гормонов тем, что их выделяют не специализированные клетки желез, а клетки тканей или отдельные самостоятельные клетки (тромбоциты, лейкоциты). Большинство факторов роста паракринны, то есть они предназначены для сигнализации на коротком расстоянии в пределах одного органа, а есть аутокринные факторы роста, которые стимулируют выделяющие их клетки. В рамках такой классификации к факторам роста следует, очевидно, относить и фитогормоны, то есть сигнальные молекулы, управля-ющие ростом и дифференциацией клеток растений. В качестве примера таких сиг-нальных молекул у теплокровных можно назвать фактор роста из тромбоцитов, кото-рый выделяется и из многих других клеток, эпидермальный фактор роста, колоний-стимулирующие факторы роста (стимулирует рост колоний некоторых лейкоцитов, поэтому их называют также интерлейкинами). Термин «цитокины» относится к факто-рам роста, участвующим в иммунных ответах.
Нейромедиаторы участвуют в работе клеток нервной ткани – нейронов.
Регуляторная система организма находится под иерархическим контролем, где основ-ная роль принадлежит нервной системе, которая таким образом через нервные окон-чания (нейромедиаторы) и поступающие в жидкие среды организма (по лат. humor – жидкость) гормоны управляет всеми функциями организма (отсюда термин нейро-гуморальная регуляция).
Рецепторные клетки кожи, слизистых оболочек, органов чувств, хеморецепторы внутри организма постоянно сообщают в ЦНС об изменениях в окружающей среде, в составе крови, в работе органов. ЦНС анализирует поступающую информацию обычно на бес-сознательном уровне и выдает адекватные команды по нервам или с помощью сигналь-ных молекул. Регуляцию с помощью сигнальных молекул осуществляет специализиро-ванный отдел головного мозга, называемый гипоталамусом. Это одна из древнейших и наиболее жизненно важных частей мозга. Гипоталамус (подбугорье) с помощью регу-ляторных молекул, называемых релизинг-факторами или либеринами, по принципу обратной связи управляет работой гипофиза – дирижера работы всей эндокринной системы организма. Гипофиз человека весит 0,5-0,6 г. Он связан с гипоталамусом короткой ножкой и состоит из трех гистологически различных частей – передней доли, промежуточной и задней доли. Передняя доля состоит из железистых клеток, работой которых управляют либерины, поступающие по кратчайшему пути из гипоталамуса по кровеносным сосудам в ножке. Задняя доля гипофиза состоит из нервных клеток, гор-моны гипоталамуса поступают сюда по нервным волокнам - аксонам.
Гормоны передней доли гипофиза, стимулирующие работу желез внутренней секреции, называются тропинами или тропными гормонами. Принцип обратной связи состоит в том, что выделяемые железами внутренней секреции гормоны тормозят выделение как либеринов, так и тропинов. Правда, здесь нет такой четкой иерархии и не все железы внутренней секреции находятся под контролем гипоталамуса при помощи либеринов. Так, например, задняя доля гипофиза, секретирующая окситоцин и вазопрессин, регу-лируется по другим механизмам, а передняя доля гипофиза секретирует самостоятель-ный гормон – соматотропин (его секреция, как и секреция других гормонов передней доли гипофиза, тормозится соматостатином – гормоном из гипоталамуса). В поджелу-дочной железе секреция инсулина (-клетки) и глюкагона (-клетки) напрямую зависит от уровня сахара в крови, но это не значит, что ЦНС не участвует в регуляции уровня глюкозы в крови. Кроме инсулина и глюкагона образование и расходование глюкозы контролируют многие другие гормоны. Под прямым контролем со стороны ЦНС нахо-дятся клетки мозгового вещества надпочечников, содержащие везикулы (пузырьки) с адреналином и норадреналином (около 20 %) и АТФ (около 4 %). По команде ЦНС в течение нескольких секунд происходит выброс адреналина за счет экзоцитоза и его концентрация в крови возрастает на три порядка (от 10-10 до 10-7 моль/л). Обычная концентрация гормонов в крови от 10-12 до 10-9 моль/л.
В качестве примера классической связи регуляции по схеме гипоталамус – гипофиз –железа внутренней секреции можно привести цепочку тиролиберин – тиреотропин – тироксин. Тиролиберин представляет собой трипептид – пироглутамил-гистидинил-пролинамид
Г ормональная
активность этого вещества поразительна.
Его содержание в гипотала-мусе настолько
мало, что для установления его строения
из 4 т гипоталамуса животных был выделен
всего лишь 1 мг этого вещества. При
введении его в организм в микро-граммовых
дозах секреция тиротропина, который
представляет собой гликопротеин с
молекулярной массой около 28 кД, возрастает
уже через несколько минут.
Метили-рование тиролиберина в
положение 1 имидазольного кольца или
замена амидной груп-пы в
пролиновом фрагменте на карбоксильную
снижает активность тиролиберина в
несколько тысяч раз, а метилирование
имидазольного кольца в положение 3 в
нес-колько тысяч раз
повышает активность. Интересно, что
этот необычный трипептид сравнительно
устойчив к действию протеаз пищеварительной
системы, поскольку он сохраняет активность
и при приеме через рот. В ответ на секрецию
тиротрипина щито-видная железа выделяет
в кровь тироксин и трииодтиронин,
образующиеся из тирози-новых фрагментов
в специально предназначенной для этого
белковой молекуле:
ормональная
активность этого вещества поразительна.
Его содержание в гипотала-мусе настолько
мало, что для установления его строения
из 4 т гипоталамуса животных был выделен
всего лишь 1 мг этого вещества. При
введении его в организм в микро-граммовых
дозах секреция тиротропина, который
представляет собой гликопротеин с
молекулярной массой около 28 кД, возрастает
уже через несколько минут.
Метили-рование тиролиберина в
положение 1 имидазольного кольца или
замена амидной груп-пы в
пролиновом фрагменте на карбоксильную
снижает активность тиролиберина в
несколько тысяч раз, а метилирование
имидазольного кольца в положение 3 в
нес-колько тысяч раз
повышает активность. Интересно, что
этот необычный трипептид сравнительно
устойчив к действию протеаз пищеварительной
системы, поскольку он сохраняет активность
и при приеме через рот. В ответ на секрецию
тиротрипина щито-видная железа выделяет
в кровь тироксин и трииодтиронин,
образующиеся из тирози-новых фрагментов
в специально предназначенной для этого
белковой молекуле:
Р егуляция
высвобождения факторов роста протекает
по многим механизмам. Они участвуют в
процессе развития, репарации поврежденных
клеток, в их дифференциа-ции, но точные
данные о всех взаимодействия на этом
уровне пока не получены.
егуляция
высвобождения факторов роста протекает
по многим механизмам. Они участвуют в
процессе развития, репарации поврежденных
клеток, в их дифференциа-ции, но точные
данные о всех взаимодействия на этом
уровне пока не получены.
По химической природе гормоны, продуцируемые железами внутренней секреции, относятся к трем типам соединений. Это могут быть пептиды, производные тирозина, а также стероиды. Растворимые в воде пептиды и гормоны на основе тирозина не про-ходят через клеточные мембраны. Их действие опосредовано рецепторными белками на внешней стороне клеток. Они, как в случае инсулина, могут работать в комплексе с транспортными белками или же действовать через специальные внутриклеточные бел-ки (G-белки) на фермент, синтезирующий внутриклеточный регулятор (его называют мессенджером). В качестве примера второго механизма действия гормонов можно наз-вать адреналин. Связываясь с рецептором он запускает механизм образования активной компоненты из G-белка (с участием ГТФ), и уже эта активная компонента включает работу аденилатциклазы, катализирующей образование ц-АМФ (это и есть мессен-джер) из АТФ. Действие G-белка ограничено по времени, так как он представляет собой гидролазу, разлагающую ГТФ на ГДФ и фосфат, а комплекс G-белка с ГДФ неактивен.
В отличие от пептидов и производных тирозина стероиды достаточно легко проходят через клеточные мембраны. Их рецепторы представлены растворенными в цитоплазме белками, которые связаны с белками теплового шока (Hsp). Обычно эти белки управля-ют организацией пространственной структуры синтезируемого на рибосоме белка (за это их называют еще шаперонами от фр. гувернантка). Гормон, проникший через мем-брану в клетку, связывается с рецепторным белком, который теперь вместе с белком теплового шока или уже без него проникает в ядро и связывается с соответствующим участком ДНК, активируя его. Однако для активации ДНК обязательно надо, чтобы белок теплового шока отошел от комплекса рецептора и гормона. Активация ДНК приводит к образованию РНК (транскрипция), РНК выходит из ядра и поступает в рибосомы, на которых уже синтезируется белок (трансляция), образование которого и должен был запустить гормон.
Из этого следует, что действие растворимых в воде гормонов проявляется в очень ко-роткие сроки (до нескольких секунд, как у адреналина), тогда как стероидные гормоны проявляют свое действие через достаточно медленно текущие процессы, в числе кото-рых транскрипция и трансляция. Действие нерастворимых в воде гормонов направлено на биосинтез белков (ферментов, транспортных белков, структурных белков), поэтому их эффект может проявиться и через несколько дней.
Из пептидных гормонов более других изучен, конечно, инсулин, а также вазопрессин, окситоцин, соматотропин, ангиотензин.
Недостаток инсулина или ослабленная реакция на инсулин со стороны клеток являются причиной сахарного диабета (в США каждый двадцатый имеет нарушения в обмене глюкозы), смертность от диабета на третьем месте. При диабете происходит серьезное отклонение от нормы в составе мочи и крови. Для усиления транспорта глюкозы в клетки у организма остается одна единственная возможность – повышение ее концентрации в крови, но почки не рассчитаны природой на этот вариант и переводят глюкозу в мочу. Кроме повышенной концентрации глюкозы (в сутки с мочой может уходить до 100 г), в моче диабетиков повышена концентрация кетоновых тел (ацето-ацетат, ацетон и -гидроксибутират), мочевины, ионов аммония (это все побочные продукты образования глюкозы из аминокислот), натрия (рН мочи у диабетиков пони-жен). Выделяющийся с выдыхаемым воздухом ацетон имеет характерный органичес-кий запах похожий на запах перегара. Диабетики много пьют, поскольку для выведения всего набора необычных отходов их жизнедеятельности расход воды увеличивается.
Антагонистом инсулина является глюкагон – тоже полипептидный гормон, выделяе-мый поджелудочной железой, но клетками другого типа. Он по действию похож на адреналин, но в отличие от него не влияет на частоту сердечных сокращений и на давление крови.
Выделяемый передней долей гипофиза самостоятельный гормон соматотропин – белок с молекулярной массой около 21 кД – регулирует вес и рост тела. Кроме того, сома-тотропин сложным образом регулирует секрецию инсулина и глюкагона, поэтому его используют для лечения некоторых форм сахарного диабета. Недостаток соматотропи-на приводит к карликовости, а избыток к гигантизму. Если усиленное выделение сома-тотропина происходит во взрослом возрасте – то это акромегалия, слоновая болезнь.
Вазопрессин повышает кровяное давление, его недостаток вызывает так называемый «несахарный диабет».


Этот гормон регулирует работу почек, он вызывает обратное всасывание воды в почках (вместе с необходимыми организму веществами). При недостатке этого гормона почки выделяют до 10 л в день сильно разбавленной мочи.
Окситоцин вызывает сокращение гладкой мускулатуры, и его применяют для стимуля-ции родов.


Тонус кровеносных сосудов регулируется ангиотензином. Этот гормон также участвует в регуляции работы почек. В неактивном виде обладающий гормональной активностью ангиотензин II содержится в составе белка ангиотензиногена. Под действием почечного ренина от ренина сначала отщепляется неактивный ангиотензин I,
![]()
от которого под действием содержащегося в плазме ангиотензин-I-превращающего фермента отщепляется две С-концевых аминокислоты и только после этого образуется активный ангиотензин II. При повышенном давлении можно получить положительный эффект за счет снижения уровня этого гормона. Во всех клетках содержатся гидролазы, расщепляюшие его, но их активация проблематична. В лекарственных целях блоки-руют превращение ангиотензина I в ангиотензин II. Для этого используют аналогичные пептидам вещества, первым из которых был каптоприл


Вторая формула принадлежит современному средству – эналаприлу. Ангиотензин действует также на надпочечник, стимулируя выделение альдостерона, что приводит к задержке в организме ионов натрия.
Гипоталамус с помощью гормона кортиколиберина через адренокортикотропный гор-мон гипофиза (АКТГ, пептид из 39 аминокислот, период полупревращения в организме около 10 мин., эффект Met-Glu-His-Phe) регулирует работу еще одной важной парной железы – надпочечников. Эти железы также состоят из двух типов секретирующих кле-ток. Клетки внешнего, коркового слоя надпочечников секретируют гормоны стероид-ного типа, которые участвуют в регуляции углеводного обмена и водно-солевого баланса – глюкокортикоиды и минералокортикоиды.

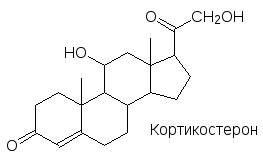

Здесь приведены три типичных представителя этих гормонов, всего их известно около 30. Все эти гормоны обладают как глюкокортикоидной, так и минералокортикоидной активностью, но у кортизола более выражена глюкокортикоидная активность, у альдо-стерона – минералокортикоидная, а кортикостерон в равной мере обладает обеими видами активности. Кортизол стимулирует образование глюкозы из аминокислот (глю-конеогенез) и обладает противовоспалительной и противоаллергической активностью. Избыток кортизола в организме приводит к потере мышечной массы, быстрой утомляе-мости, при общей худобе лицо у человека становится пухлым, круглым (болезнь Кушинга). Минералокортикоиды вызывают задержку ионов натрия в организме и поте-рю ионов калия с мочой. Недостаточная секреция гормонов коры надпочечников при-водит к аддисоновой болезни, характеризующейся слабостью, подверженностью к стрессам и к инфекциям, кожа приобретает характерный цвет (бронзовая болезнь).
Лекарственное применение кортикостероидов осложнено нарушением гормонального баланса организма из-за реализации принципа обратной связи (их длительное введение в виде лекарства может привести к атрофии надпочечников). Сейчас в основном стара-ются использовать их в виде средств местного действия и для этого уменьшают их растворимость и сродство к транспортным белкам за счет введения атомов фтора и гидрофобизации


Еще одна группа стероидных гормонов представлена половыми гормонами (андрогены, эстрогены и гестагены). Кора надпочечников, семенники и яичники имеют общее эмбриональное происхождение, поэтому в разных количествах мужские и женские половые гормоны выделяются всеми этими типами тканей. То есть их действие зависит от соотношения между ними.

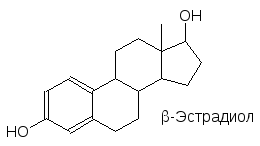

Уровень этих гормонов также контролируется гипоталамусом и гипофизом через лютеинизирующий гормон и гонадотропины.
Половые гормоны определяют внешние половые признаки – мужские (андрогены) или соответственно женские, они регулируют процессы роста, поддерживают функцио-нирование продуктивной системы. Андрогены стимулируют рост скелетных мышц и поэтому их часто называют анаболическими гормонами. Эти свойства привлекают вни-мание фармакологов, поскольку разделение анаболической и андрогенной активности перспективно не только с целью получения анаболических допингов для спортсменов, но и с фармакологической точки зрения для лечения некоторых видов мышечных сла-бостей (астенические синдромы).
Э ффективным
анаболиком оказался 19-нортестостерон,
который в виде его эфира с фенилпропионовой
кислотой используется на практике под
названием феноболин
ффективным
анаболиком оказался 19-нортестостерон,
который в виде его эфира с фенилпропионовой
кислотой используется на практике под
названием феноболин
Тестостерон и особенно метилтестостерон задерживают развитие рака молочных желез, но побочный андрогенный эффект этих гормонов не позволяет использовать их для лечения этого заболевания.
Точно также женский половой гормон эстрадиол был бы неплохим средством для лече-ния атеросклероза, но и здесь связанное с ним появление женских вторичных половых признаков не позволяет использовать его для этих целей. Тем не менее женские поло-вые гормоны используются в медицинской практике для восполнения их недостатка после удаления яичников или в менопаузе. Но наиболее широкое применение нашли они в качестве пероральных контрацептивов. В этом случае экзогенные гормоны не вызывают интенсивной перестройки гормональной системы или атрофии генерирую-щих их тканей (как это было отмечено в случае кортикостероидов), так как периоди-ческие колебания их концентрации в крови характерны для женского организма. Наиболее эффективны сочетания эстрогенов и гестагенов (гормонов желтого тела). Здесь были приложены колоссальные усилия по поиску оптимальных структур анало-гов и их дозировки. В качестве эстрогена в составе этих средств чаще всего исполь-зуется этинилэстрадиол, а в качестве гестагена – прегнин


Принцип действия этих средств основан на создании гормональной среды, характерной для состояния беременности. Все эти средства принимаются при врачебном контроле, а современные препараты вообще состоят из двух или трех различных таблеток – эстро-гена, гестагена и, желательно, гормонов гипофиза и гипоталамуса. Причем их дозы подбираются лечащим врачом с учетом гормонального статуса принимающей их женщины.
В общем случае надо представлять себе, что нормальный предусмотренный природой жизненный цикл женщины представляет собой чередование периода беременности и кормления ребенка грудью без, если можно так сказать, простоев. Иначе говоря, как раз менструация – это ненормальность (недаром она так болезненна). Регулирующий лак-тацию и, как это всегда бывает у гормонов, многие другие функции, включая прояв-ление материнского рефлекса, гипофизарный гормон пролактин (белок) в период беременности и активного кормления грудью (его концентрация при этом возрастает от 5-10 до 200 нг/мл) препятствует овуляции, именно поэтому раньше длительное, до двух-трех лет, кормление ребенка грудью было и остается лучшим естественным противозачаточным средством. Но его применение в качестве экзогенного противоза-чаточного средства связано с трудностью равномерного введения высокомолеку-лярного белка (23-24 кДа) и с активизацией им процесса лактации.
Гормоны регулируют также рост и развитие насекомых. Более всего у этих предста-вителей животного мира изучены гормоны линьки. Сам процесс линьки запускается гормоном стероидной природы – экдизоном

Н о
высшие насекомые, проходящие в своем
развитие несколько фаз (яйцо, личинка,
куколка, имаго), претерпевают так
называемый метаморфоз – переход из
одной фазы развития в другую. А это уже
регулируется ювенильными гормонами:
при их наличии линька не сопровождается
метаморфозом. Приведена структура
ЮГ-III:
о
высшие насекомые, проходящие в своем
развитие несколько фаз (яйцо, личинка,
куколка, имаго), претерпевают так
называемый метаморфоз – переход из
одной фазы развития в другую. А это уже
регулируется ювенильными гормонами:
при их наличии линька не сопровождается
метаморфозом. Приведена структура
ЮГ-III:
Образование ювенильных гормонов блокируется, например, фтормевалонатом,

а агонистом ювеноидов являются очень многие вещества, например метопрен

При действии фтормевалоната личинка преждевременно претерпевает метаморфоз и превращается в неполноценную, как бы карликовую форму взрослого насекомого, которое не может дать потомства, а действие метопрена приводит к тому, что личинка не превращается во взрослое насекомое и также не может дать потомства. Это уже ин-сектициды четвертого поколения, к ним-то и термин инсектициды не очень подходит.
Особое место занимают фитогормоны, вещества регулирующие рост и развитие расте-ний, но в приведенной выше классификации эти вещества скорее всего надо относить к факторам роста, поскольку их образование идет не в специализированных клетках (клетках желез), а в клетках меристемы (это растительный аналог эмбриональной ткани), в клетках корней, листьев, то есть в обычных составляющих растение тканях. Здесь также очень важную роль играет сочетание различных концентраций гормонов и их взаимное влияние. Так, например, повышенная концентрация гормона роста ауксина приводит к образованию ингибитора роста – этилена. Разные концентрации двух гор-монов роста – ауксина и цитокинина – определяют стимуляцию роста и дифферен-циации различных клеток.