

План:
I. Общие сведения
IIХарактеристики вызванных потенциалов
IIIКлассификация вызванных потенциалов
IVСхема оборудования
VЭлектроды
VI Анализ вызванных потенциалов. Метод вычисления мгновенной мощности.
VII Математический анализ вп.
VIII Заключение.
IX. Список используемой литературы и ссылки в Интернете.
I.
Одним из основных методов анализа деятельности мозга является изучение биоэлектрической активности различных структур, сопоставление записей, одновременно отводимых от различных участков мозга, как в случае спонтанной активности этих структур, так и в случае электрических реакций на кратковременные одиночные и ритмические афферентные стимулы. Часто используются также одиночное или ритмическое электрическое раздражение тех или иных образований мозга с записью реакций в других структурах.
Метод вызванные потенциалы (ВП) давно является одним из ведущих в экспериментальной нейрофизиологии; с помощью этого метода получены убедительные данные, раскрывающие сущность ряда важнейших механизмов мозга. Можно с уверенностью считать, что большая часть сведений о функциональной организации нервной системы получены с помощью этого метода. Развитие методов, позволяющих записывать ВП у человека, открывает блестящие перспективы для изучения психических заболеваний.
Регистрация ответов нервов и отдельных нервных волокон на электрические стимулы позволила исследовать основные закономерности возникновения и проведения нервных импульсов в нервных проводниках. Анализ ответов отдельных нейронов и их скоплений на раздражение выявил основные законы возникновения торможения и возбуждения в нервной системе. Метод ВП является главным способом установления наличия функциональных связей периферии с центральными нервными механизмами и исследования межцентральных соотношений в нервной системе. Регистрируя ВП, удалось установить основные закономерности функционирования специфической и неспецифической систем афферентации и их взаимодействия между собой.
Методом ВП изучены характеристики изменения реактивности ЦНС на афферентные стимулы в зависимости от уровня функциональной активности мозга; исследованы закономерности взаимодействия синхронизующих и десинхронизующих систем ствола, таламуса и переднего мозга.
Исследования ВП различных уровней нервной системы являются основным методом тестирования действия фармакологических нейротропных препаратов. С помощью метода ВП успешно изучают в экспериментах процессы высшей нервной деятельности: выработку условных рефлексов, сложные формы обучения, эмоциональные реакции, процессы принятия решения.
Методика ВП прежде всего применима для объективного тестирования сенсорных функций (зрения, слуха, соматической чувствительности), для получения более точных сведений о локализации органических церебральных поражений, для изучения состояния проводящих путей мозга и реактивности различных церебральных систем при патологических процессах.
Наиболее широкое применение в качестве метода оценки состояния сенсорной системы исследование ВП нашло в области изучения нарушений слуховой функции; методика получила название объективной аудиометрии. Преимущества ее очевидны: появляется возможность исследовать слух у детей грудного возраста, у лиц с нарушением сознания и контакта с окружающими, в случаях истерической и симулируемой глухоты. Также, путем регистрации ВП от брюшной стенки матери в области, соответствующей головке плода, можно выявлять степень развития функций слуха у плодов человека.
Достаточно перспективным представляется изучение зрительных ВП (ЗВП), учитывая большое значение оценки состояния зрительных систем в топической диагностике церебральных поражений.
Исследование соматосенсорных ВП (ССВП) позволяет определять состояние сенсорных проводников на всем протяжении от периферии до коры. Поскольку ССВП имеют соматотопику, соответствующую корковым проекциям тела, особый интерес приобретает их исследование при поражении сенсорных систем на уровне головного мозга. Большое практическое значение может иметь исследование ВП с целью дифференциации органических и функциональных (невротических) сенсорных нарушений. Это дает основание использовать методику ССВП в судебной медицине.
Большой интерес представляет исследование ВП при эпилепсии, учитывая большую роль, которую играет афферентная импульсация в патогенезе развития эпилептических припадков. Высокая чувствительность ВП к изменениям функционального состояния мозга под влиянием фармакологических препаратов позволяет использовать их с целью тестирования эффектов лечения при эпилепсии.
Помимо исследования ВП на относительно простые стимулы (короткая вспышка света, звуковой щелчок, короткий импульс электрического тока), в последнее время появился ряд исследований ВП на более сложные виды стимуляции с использованием также более сложных способов выделения и анализа ВП. В частности, достаточно широко исследуются ВП на предъявление зрительных стимулов, представляющих собой изображение. Чаще всего используют изображение синусоидальную модулированной по яркости или контрастной решетки или шахматного рисунка с различными пространственными частотами и мерой контраста. Изображение предъявляют как относительно продолжительным засвечиванием. Кроме того, используют предъявление с помощью синусоидально модулированного во времени по яркости светового потока. Пользуясь этим методом, получают так называемые ВП постоянного состояния. Этот ВП представляет собой колебательный синусоидальный процесс с постоянными частотно-амплитудными характеристиками, находящийся в определенном частотно-амплитудном соотношении с частотой и интенсивностью светового потока, осуществляющего визуальную стимуляцию. Такие потенциалы чаще всего применяют в тестированиях функции зрения, причем в настоящее время исследования не выходят в основном за рамки лабораторных экспериментов.
Существенное практическое значение в клинических исследованиях приобретают ВП на извращения зрительного паттерна (когда черные элементы на экране меняются местами с белыми). Получены данные, показывающие закономерную связь амплитуды и латентных периодов некоторых компонентов этих ВП с размерами шахматного поля и корреляцию с остротой зрения. С точки зрения клинической неврологии наибольший интерес представляют ВП на извращение зрительного паттерна в исследованиях демиелинизирующих заболеваний.
В последние годы были проведен анализ как ВП в норме с точки зрения их связи с различными звеньями афферентных систем, так и исследование изменений ВП при патологии с точки зрения связи этих изменений с общими и частными перестройками возникающими в ЦНС под влиянием патологического процесса.
Исследование ВП находят применение во многих областях клинической практики:
-
локальное деструктивные поражения нервной системы:
-
поражения периферической нервной системы;
-
поражение спинного мозга;
-
поражение ствола мозга;
-
поражение полушарий мозга;
-
поражение таламуса;
-
супраталамические поражения;
-
нервные болезни:
-
эпилепсия;
-
опухали ЦНС;
-
церебрально-сосудистые нарушения;
-
черепно-мозговая травма;
-
деминиции;
-
метаболические нарушения;
-
кома и вегетативное состояние;
-
реанимационный мониторинг [6].
Возможности метода ВП позволяют не только обнаружить структурный уровень поражения анализатора, но и количественно оценить характер поражения сенсорной функции человека в различных звеньях анализатора. Особую ценность и уникальность метод регистрации ВП представляет для обнаружения сенсорных нарушений у детей самого раннего возраста. Системы использующие метод ВП используются в нейрологии, нейрохирургии, дефектологии, клинической аудиометрии, психиатрии, судебно-психиатрической, военной и трудовой экспертизе [3].
II. Вызванные потенциалы коры, или вызванные ответами, называют градуальные электрические реакции коры на однократное афферентное раздражение какого-либо раздела нервной системы. Амплитуда, которых в норме достигает 15 мкВ – длинолатентные (до 400 мс) и 1мкВ – коротко латентные (до 15 мс) [2].
Если бы ВП были связаны только с одним определенным типом активности нейронов, количественная оценка их была бы сравнительно легкой, так как сама кривая ВП представляла бы отражение этой активности. Однако гораздо более вероятно, что ВП генерируются несколькими типами активности с различными временными и, возможно, пространственными характеристиками. Помимо других данных, об этом свидетельствует сложность волновой конфигурации ВП, представленная наличием множества пиков, появляющихся через различные временные интервалы. В соответствии с этим исследователи говорят о различных компонентах, определяемых на основании латентности и полярности видимых максимумов и минимумов (пиков) на кривой. При количественной оценке важно определить, на каком протяжении будут измеряться избранные компоненты.
|
|
Рис.1. Модель, образования ВП. |

На рис.1 показана типичная модель,
позволяющая понять как образуются ВП.
Отчетливо виден, (рис.1А) ряд позитивных
негативных колебаний, каждое из которых
наступает через определенный промежуток
времени, но их временные параметры
частично совпадают. Таким образом, волны
a![]() и в кривой представляют собой
негативные колебания, разделенные во
времени, а волны б и г - позитивные
колебания, перекрывающиеся во времени.
На рис. 1Б показана кривая, полученная
при суммации волн
а, б, в
и г.
Отчетливо видно, что появляющийся
негативный компонент в не отражает
истинной негативности кривой. Более
того, максимальная активность (компонент
б и г) смещена во времени, поэтому
максимумы этих компонентов, определяемые
на основании их
латентности, не являются истинными
максимумами. Правильность модели,
изображенной на рис.1А, подтверждается
косвенными данными .
и в кривой представляют собой
негативные колебания, разделенные во
времени, а волны б и г - позитивные
колебания, перекрывающиеся во времени.
На рис. 1Б показана кривая, полученная
при суммации волн
а, б, в
и г.
Отчетливо видно, что появляющийся
негативный компонент в не отражает
истинной негативности кривой. Более
того, максимальная активность (компонент
б и г) смещена во времени, поэтому
максимумы этих компонентов, определяемые
на основании их
латентности, не являются истинными
максимумами. Правильность модели,
изображенной на рис.1А, подтверждается
косвенными данными .
III Воган различает 5 классов таких потенциалов:
1. Сенсорные ВП
2. Моторные ВП
3. Потенциалы с большим латентным периодом, связанные со сложными
психологическими факторами
4. Сдвиги постоянного потенциала
5. Потенциалы немозгового происхождения
Сенсорные ВП в свою очередь подразделяются в зависимости от мо-
дальности на зрительные, соматосенсорные, слуховые и в зависимости от
периода времени анализа (иначе говоря времени возникновения потенциа-
ла после стимула) на коротколатентные (мозгового ствола), средне- и
длиннолатентные.
Зрительные вызванные потенциалы
При использовании соответствующих зрительных раздражителей в зрительных путях может быть вызван синхронный импульс, что ведет к возникновению корковых потенциалов с определенной формой волны и определенным периодом латентности. Чаще всего используются такие зрительные раздражители, как повторные вспышки света и/или обращение шахматного паттерна. Кортикальные реакции исследуются с помощью поверхностных электродов, располагающихся над затылочной областью. Наиболее выраженным и постоянным ответом является компонент Р100, встречающийся практически у всех и в норме имеющий латентный период от 90 до 120 мс, однако в большинстве случаев укладывающийся по пиковой латентности в достаточно узком диапазоне в области 100 мс. Зрительные вызванные потенциалы позволяют оценить функциональное состояние зрительного анализатора от зрительного нерва до зрительной коры.
Показания:
- Неврит зрительного нерва или травматическое его поражение; - Демиелинизирующие заболевания; - Корковая слепота и другие повреждения зрительной коры; - Психогенная слепота у истерических больных - И т.д.
Противопоказания - при стимуляции на реверсивный шахматный паттерн: - Дети до 3 лет; - Лица с остротой зрения менее 0,1; - Отсутствие рефракции или фиксация взора у пациента; - Отсутствие оптической коррекции; - Применение препаратов, расширяющих зрачок, за 2-3 суток до обследования. Специальной подготовки к исследованию не требуется.
Соматосенсорные вызванные потенциалы
Это электрические ответы нервных волокон и центров на стимуляцию рецепторов соматической чувствительности или соответствующих им нервных стволов. В практической клинической диагностике применяют только электрическую стимуляцию. Регистрирующие электроды располагают вдоль нервных стволов, в области сплетений, спинальных центров. Кортикальный потенциал регистрируется над контралатеральной сенсорной корой. Исследование соматосенсорных ВП позволяет определить состояние сенсорных проводников на всем протяжении от периферии до коры.
Показания:
- Невропатии, поражение корешков; - Поражение плечевого сплетения; - Поражения различных отделов спинного мозга; - Рассеянный склероз; - Оценка нарушений сенсорных функций у больных истерией; - Различные патологические состояния, приводящие к поражению на уровне ствол - сенсорная зона коры; - Определение тяжести повреждения мозга и возможных последствий травмы и комы; - Диагностика смерти мозга. Специальной подготовки к исследованию не требуется.
Акустические стволовые вызванные потенциалы
Регистрация ответов, вызванных акустической стимуляцией, соответствующих электрическим реакциям на раздражение кохлеарных ядер, а также путей и ядер нижнего отдела мозгового ствола. При моноаурикулярной стимуляции обычно используют щелкающий звук с частотой 10 Гц, интенсивность звука устанавливается на 65 - 70 дБ выше порога слышимости; для другого уха используется маскирующий шум. Обычно суммируется реакция на 1000 - 2000 щелчков. Стволовые потенциалы обозначаются римскими цифрами I - VII. Определяются, в частности, межпиковые латентные периоды и соотношение амплитуд волн I/V. Волна I представляет собой потенциал действия слухового нерва, волна II происходит из кохлеарного ядра, волна III - из верхнего оливарного; волна IV связана с латеральной петлей, V - с нижним бугорком; VI - с медиальным коленчатым телом и VII - с таламокортикальными проекциями, включая, возможно, первичный ответ коры. Волны VI и VII отличаются непостоянством и вариабельностью. В клинической неврологии метод акустических стволовых ВП применяется для оценки целостности периферических и стволовых невральных структур, участвующих в проведении и восприятии слуховой информации, а также для уточнения возможного уровня их поражения.
Показания:
- Нейросенсорная тугоухость; - Невринома VIII пары; - Демиелинизирующие заболевания; - Оценка сохранности слуха у детей раннего возраста; - Оценка состояния больных с комой, прогноз выхода из нее; - Диагностика смерти мозга.
IV.
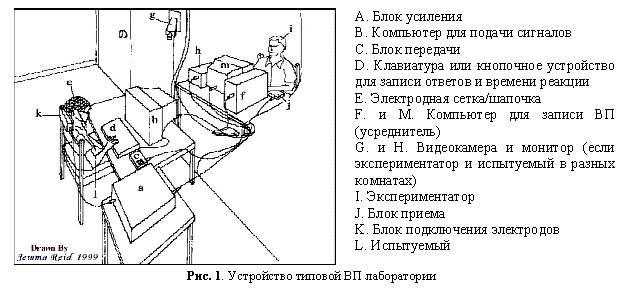
Лабораторное оборудование для записи ВП представляет собой систему, состоящую из нескольких компонентов. На рисунке 1 изображена типовая кофигурация такой системы.
V
Главным компонентом ВП системы является электродная шапочка (см. Е на Рис. 1). Количество электродов, иначе еще называемых каналами (channels), обычно кратно 32, поэтому минимальное количество электродов тоже 32. Более распространенными являются шапочки с 64, 128 и 256 электродами. Системы, использующие 128 или 256 электродов, называются высокоплотнымы (high density). При психолингвистических исследованиях в основном используются системы с 64 или 128 электродами.
Перед началом процедуры установки электродов замеряется размер головы испытуемого, так как электроды крепятся к специальной эластичной шапочке. Размеры шапочек для взрослых испытуемых колеблются от 48 до 60 см, в зависимости от окружности головы. Существуют специальные шапочки для младенцев (31-42 см) и детей дошкольного возраста (42-48 см). Шапочки бывают двух видов (Рис. 2): шапочка-сетка, с неснимающимися электродами (компания EGI) и шапочка со вставляющимися электродами (компания Neuroscan). Шапочка-сетка с неснимающимися электродами (Рис. 2а) определенным образом одевается на голову испытуемого и не требует никаких специальных операций для ее подготовки.


При использовании шапочки со вставляющимися электродами процедура подготовки испытуемого к эксперименту занимает от 30 минут до часа. В шапочке проделаны отверстия по количеству электродов, в которые вставлены пластмассовые колечки-держатели. До начала эксперимента лаборант подготавливает шапочку, а именно, вставляет каждый электрод в пластмассовые колечки на шапочке.

Каждый электрод состоит из металлического кольца на одном конце и штепселя на другом (Рис. 3). Металлическое кольцо обычно сделано из специального серебряного сплава (AgAgCl) и вставляется в пластмассовое колечко-держатель на шапочке. Каждый электрод имеет свое обозначение и прикрепляется в строго определенное место на шапочке. Сопротивляемость электродов обычно устанавливается ниже 5 кW. Эксперименты на материале русского языка, о которых пойдет речь в разделе 6, проводились при помощи ВП системы Нейроскэн, которая используется для психолингвистических экспериментов, проводимых в институте Лингвистики университета Потсдама в Германии (руководители лаборатории профессора Даглас Сэдди, Маттиас Шлезевский и научный сотрудник Штефан Фриш) [31]. Дальнейшее описание методики ВП будет на примере этой системы.
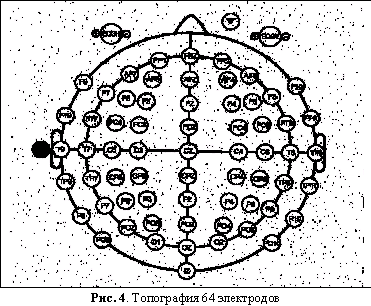
На Рис. 4 изображена типовая топография ВП системы с 64 электродами. Помимо 64 электродов на голове используются еще два референциальных электрода (не изображенных на Рис. 4) и четыре глазных электрода (EOGH и EOGV). Референциальные электроды, называемые мастоидными (mastoids), устанавливаются на черепные кости, расположенные непосредственно за мочками ушей (на иллюстрации 4 изображен в виде черного кружка рядом с левым ухом). Референциальные электроды являются точками отсчета: электроэнцефалограмма вычисляется при помощи вычитания потенциалов, записанных на всех остальных точках головы, из потенциалов, записанных в точках отсчета. Четыре глазных электрода устанавливаются на лице, по одному у уголков глаз, и два на правый глаз, один сверху, другой снизу (см. Рис. 2). Глазные электроды записывают электроокулограмму (electrooculogram, EOG), т.е., вертикальные и горизонтальные движения глаз, которые впоследствии отфильтровываются.
Головные электроды делятся на несколько групп, в зависимости от их расположения: на левой или правой стороне головы и, соответственно, на левом или правом полушарии и в разных долях. Электроды левого полушария имеют нечетные номера 1-9, правого – четные номера 2-10, при этом номера возрастают от центра к периферии. В соответствии с долями электроды делятся на следующие группы:
А – передние доли (anterior)
P – задние доли (posterior)
Z – средняя линия (midline)
F – лобная (frontal)
T – височная (temporal)
C – центральная (central)
P – теменная (parietal)
O – затылочная (occipital).
А и Р участвуют в обозначении только в комбинации с Z, F, Т и O. Так, электрод CZ располагается точно в самой середине головы, рядом с ним находится электрод заземления (на Рис. 4 не обозначен), T10 – височный электрод непосредственно над правым ухом, F3 – лобный электрод, расположенный посередине левой лобной доли, PO8 – затылочный электрод, находящийся сзади и в самом низу правого полушария. Количество электродов, задействованных в эксперименте, может варьироваться в каждом конкретном случае.
VI
Традиционные методы обработки любых длиннолатентных вызванных потенциалов представляют собой разметку и определение положения позитивных и негативных пиков, картирование, спектральный, корреляционный анализ и некоторые дополнительные виды оценки сигнала ВП. Все они по определению требуют незашумленных , т.е. «чистых» исходных данных. В действительности же коэффициент сигнал/шум в любом зарегистрированном ВП достаточно мал и определяется соотношением уровней фоновой ритмики и собственного отклика мозговых структур на стимул, а также корнем квадратным из числа усреднений.
В клинике и в большинстве научных исследований сигнал вызванных потенциалов чаще всего анализируется без учета шумового вклада фоновой электроэнцефалограммы. При этом исследователи по умолчанию еще и принимают на веру утверждение о значительной изменчивости ВП как внутри популяции, так и у одного и того же испытуемого в различные моменты времени или при различных условиях регистрации.
Приведем упрощенный расчет соотношения сигнал/шум для типичного случая регистрации длиннолатентных ВП. Допустим, уровень фоновой электроэнцефалограммы в процессе регистрации составлял 25 мкВ от пика до пика. Уровень истинного отклика мозговых структур предположим в пределах 5 мкВ от пика до пика. Введем общепринятое допущение о полном отсутствии корреляции между двумя процессами и рассчитаем вклад фоновой ЭЭГ в зарегистрированный вызванный потенциал. Предельно упрощая и исключая специальные термины, можно сказать, что уровень электроэнцефалограммы будет уменьшаться пропорционально корню квадратному из количества поданных стимулов. Т.е. если, допустим, было подано 100 вспышек при регистрации зрительного ВП, то уровень шума в полученной смеси будет равен 25/√100=2.5мкВ от пика до пика. При предполагаемом уровне ВП равном 5 мкВ, соотношение сигнал шум в данном эксперименте будет равно 2:1, т.е. уровень помехи будет равен половине уровня истинного ВП. При подаче 400 стимулов отношение улучшится до 4:1 и т.д., но в любом случае в зарегистрированном вызванном потенциале всегда есть значительный вклад компонентов, которые не имеют никакого отношения к истинному отклику мозговых структур. Все эти расчеты подробно описаны в специальной литературе. При практической работе всегда можно с уверенностью утверждать, что не менее 10% «шумового» вклада в анализируемый вызванный потенциал обеспечивает фоновая ритмика мозга.
В то же время существующие методы анализа ВП требуют нахождения с высокой точностью вершин позитивных и негативных пиков и точное определение их временных и амплитудных показателей. На основании полученных данных и делается заключение. При этом, повторим, погрешность, обусловленная вкладом фоновой ритмики в подавляющем большинстве случаев никак не учитывается. Для случая здорового испытуемого это не так важно, потому, что фоновая электроэнцефалограмма примерно идентична в обоих полушариях. Зарегистрированные ВП будут искажены, но искажены идентично справа и слева. И только при повторной записи того же испытуемого исследователь обнаружит, что форма вызванных потенциалов изменилась, но тоже идентично справа и слева. Учитывая общепринятое мнение о существенной изменчивости формы ВП, обнаруженный феномен именно так и будет оценен. Однако при записи пациента с локальной патологией или выраженной межполушарной асимметрией вклад фоновой ритмики существенно снизит ценность и точность исследования, чего не заметит исследователь.
Описанные сложности разделения истинного отклика мозговых структур на поданный стимул и шума, обусловленного фоновой ЭЭГ, определяются самим способом регистрации ВП, т.е. корреляционным приемом. И хотя такой метод регистрации теоретически позволяет получить любое сколь угодно хорошее разделение, в действительности все ограничивается предельным числом поданных стимулов, которое не сможет быть бесконечным и не может превысить величину, при которой начинается существенное уменьшение отклика мозговых структур на подаваемые стимулы. В практическом плане это означает, что получить «чистые» ВП при реальной экспериментальной работе невозможно.
Применение экзотических способов ( ICA и др.) дополнительного разделения компонентов следует считать не вышедшими из стадии экспериментов и весьма опасными в смысле дезинформации исследователя (если выдвинутой авторами метода модель не соответствует внутренней блок-схеме мозговых структур). Что касается общепринятого метода оценки качества регистрации ВП методом вычисления корреляции между группами четных и нечетных усреднений, то следует учитывать, что величина, описывающая полученное качество регистрации является верхним теоретически возможным пределом. Реальное соотношение сигнал/шум хуже, что обусловлено коррелированными со стимулами компонентами ЭЭГ и другими, в том числе и статистическими особенностями электроэнцефалограммы.
Учитывая специфику анализа вызванных потенциалов – необходимость довольно точного измерения степени задержки распространения импульсов (в том числе и локальной) и требование обнаружения изменений амплитуды (в том числе и локальных), можно утверждать, что наличие шума в сигнале ВП существенно затрудняет работу и снижает точность метода. Кроме того, от шума страдает и интерпретация «врачебного» уровня т.к. некоторые пики могут быть смещены, искажены или даже вообще не обнаруживаться. В связи с этим несомненно, что любой способ, который может помочь повысить точность «разметки» сигнала ВП может быть чрезвычайно полезным не только в научной, но и клинической практике.
Очевидно также (доказательства опускаются), что никакой математический метод, не использующий априорных допущений или искусственных моделей, не может быть применен для улучшения соотношения сигнал/шум вызванных потенциалов. Действительно – отфильтровать шум невозможно т.к. он находится в той же частотной полосе, что и полезный сигнал, временное разделение после корреляционного приема невозможно в принципе, а вводить умозрительную модель недопустимо до тех пор, пока мы не будем знать алгоритма функционирования мозга хотя бы в области генерации ЭЭГ и ВП. В итоге остается единственная возможность – пространственная фильтрация, использующая многоканальную регистрацию сигналов.
Следует сразу оговориться, что линейное суммирование сигналов ВП от разных участков коры бессмысленно вследствие фазового несовпадения их компонентов, амплитудных различий и особенностей системы отведений (биполярной, униполярной и т.п.). Кроме того, получаемый сигнал будет иметь весьма неопределенный физический смысл. Более того, он будет вариабельным и уникальным для каждой записи каждого пациента, а для некоторых систем отведений - вырождаться в изолинию.
В связи с этим предлагается использовать суммарную мгновенную мощность вызванных потенциалов, зарегистрированных одновременно во всех отведениях. Такой параметр будет иметь вполне определенный физический смысл – это мгновенная электрическая мощность, генерируемая всеми нейронами, которые в данный момент заняты обработкой информации, полученной от внешнего стимула.
На Рис.1 представлены результаты такой обработки. Для стандартных длиннолатентных ЗВП (100 стимулов) вычислена мгновенная мощность, график которой представлен в правой части рисунка. Отдельно произведено вычисление для правого и левого полушария раздельно – графики внизу.

Рис.1 Длиннолатентные ЗВП и графики их мгновенной мощности.
Обратим внимание, что графики получились весьма информативными: в момент стимула мощность близка к нулю, отчетливо видны ранние компоненты, основной отклик, поздний колебательный процесс и затухание вызванного потенциала. Наглядно представляются асимметрии – ранние компоненты более выражены в левополушарных областях, а основной отклик имеет преимущественно правостороннюю локализацию.
Если оценивать график мощности по критерию улучшения коэффициента сигнал/шум, то необходимо учитывать, что этот сигнал имеет выигрыш перед стандартным представлением ВП в количество раз, равное корню квадратному из числа каналов записи (в действительности больше, что связано с фазовыми смещениями электроэнцефалограммы в различных отведениях). В представленном случае – не менее чем в четыре раза, т.к. использовался 16-канальный электроэнцефалограф. Разумеется, четырехкратное улучшение отношения сигнал/шум может дать принципиально новое качество точности оценки временных параметров ВП, а также обнаружения и распознавания анализируемых пиков.
Следует отметить, что график мощности нормирован, т.е. максимальное зарегистрированное значение принято за 100 процентов, а остальные выражены в долях от него. Физический смысл такой нормировки очевиден: если считать, что в момент наибольшей мощности активно максимальное для обработки мозгом подаваемого зрительного стимула количество нейронов, то в остальные моменты времени можно узнать процентное соотношение числа активных нейронов.
Очевидно также, что сопоставление графика мощности с вызванным потенциалом, зарегистрированном в любом отведении, может обеспечить достоверную временную «привязку» при определении пиков, интересующих исследователя.
Попробуем привести примеры определения пиков мощности ЗВП. Для здоровых испытуемых (Рис.2) типичным (для условий регистрации в нашей лаборатории) является форма кривой с подъемом в областях 120, 200, 270 мсек.

Рис.2 Типичная форма графика мощности для ЗВП здорового добровольца
Для пациентов с зарегистрированной пароксизмальной активностью и клиническими проявлениями эпилепсии форма графика мощности существенно меняется. Обращают внимание либо дополнительные пики в областях после 300 мсек. (Рис.3, Рис.4), либо появление дополнительного «острого» пика в областях 70 – 100 мсек. (Рис.5).

Рис.3 Изменение формы графика у пациентов с пароксизмальной активностью

Рис.4 Выраженный дополнительный пик мощности в области 400 мсек, существенное искажение формы ВП, вызывающее затруднение при «классической» разметке.

Рис.5 Появление «дополнительного» острого пика на графике мощности у пациента с пароксизмальной активностью.
Весьма важно и то, что с учетом улучшения соотношения сигнал/шум, график мощности вызванных потенциалов может быть использован для сопоставления электрофизиологических и клинических данных. Например: хорошо известный типичный феномен трансформации ВП в комплекс «пик-волна» является следствием далеко зашедшего и явно выраженного процесса. Но не исключено, что его более ранние стадии могут быть выявлены с помощью метода, предоставляющего более чистый и достоверный сигнал, чем классические ВП. Тем более вероятна связь получаемой формы графика мощности ВП с генетически детерминированными скоростями проведения нервного импульса, типами обработки информации и т.д. Не исключено также, что некоторые формы эпилепсии будут давать патогномоничные особенности графика мощности. Вообще, следует предполагать, что те особенности ВП при различных заболеваниях, которые не были выявлены исследователями вследствие физических причин, легко могут быть обнаружены при анализе графика мощности. Обоснование этого тезиса очевидно – использование для анализа менее зашумленного сигнала ведет к увеличению разрешения метода.
Можно ожидать также, что график мощности ВП проявит большую, чем обычные вызванные потенциалы, специфичность к подаваемым стимулам. При этом его форма и расположение пиков будут более стабильными и распознаваемыми. Экспериментальное подтверждение этого тезиса следует из иллюстраций.
Для экспериментальной работы использовалась записи здоровых добровольцев, у которых последовательно после промежутка для отдыха регистрировались различные типы длиннолатентных вызванных потенциалов. Анализировались собственно ВП и график их мгновенной мощности. Результаты представлены на следующих иллюстрациях.

Рис.6 Длиннолатентные ЗВП, (200 стимулов) и их графики мощности. Фотостимулятор светодиодный, длительность вспышки 8 мсек. Стимуляция симметрично на оба глаза, глаза закрыты.
В данном случае можно наблюдать хорошее совпадение формы графика мощности ВП с предыдущими наблюдениями у здоровых добровольцев. Следует отметить заметную асимметрию, особенно в области ранних компонентов и пика в 100 мсек. Основные пики графика мощности расположены на 100, 175, 270 мсек.
Очевидно, что при исследовании ВП на обращение шахматного паттерна мы должны получить другую форму вызванных потенциалов у того же испытуемого. Следует ожидать также изменения вида графика мощности. С учетом специфического действия шахматного паттерна можно ожидать смешения пиков мощности во времени и изменения самой структуры отклика мозговых структур на предъявленные стимулы.
На рис.7 наблюдается именно такая картина. Основные пики графика мощности сместились в области 90, 135, 220 мсек, уменьшилась межполушарная асимметрия, максимальный по амплитуде отклик стал более коротким т.е. кратковременным, появились дополнительные компоненты в областях 400-500 мсек.

Рис.7 Реакция на обращение шахматного паттерна (100 стимулов). Тот же испытуемый. ЗВП и график мощности существенно отличаются от предыдущей записи.
Если сравнивать ВП на вспышку и ВП на шахматный паттерн, то сами вызванные потенциалы сопоставить и проанализировать непросто, а график мощности весьма удобен для анализа наблюдаемых изменений (рис.8)

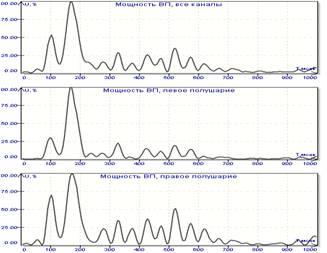
Рис.8 Сопоставление графиков мощности при регистрации ЗВП на вспышку (слева) и на обращение шахматного паттерна (справа).
При регистрации графика мощности ЗВП на изменение изображений отмечается смещение пиков в области 150, 260, 410 мсек. Увеличение латентных периодов объясняется, видимо, большей сложностью стимула и, как следствие, более длительной реакцией на него (Рис.9). Двухфазные ранние компоненты, как следует из анализа графика мощности, обусловлены, вероятно, процессом обнаружения и распознавания самого факта изменения изображения.

Рис.9 Длиннолатентные ЗВП на стимуляцию изменением изображений
Сопоставление графиков мощности ЗВП на вспышку и ЗВП на изменение изображения наглядно демонстрируют разницу ответов мозговых структур на эти стимулы (Рис.10).


Рис.10 График мощности ЗВП на вспышку света (вверху) и график мощности ЗВП на изменение изображения (внизу). Обращает внимание дополнительный (второй) ранний компонент.
Представляет интерес также обнаружение с помощью графика мощности компонентов ЗВП, связанных с когнитивными функциями. Вычислим этот график для ЗВП на смену изображений при коэффициенте усреднения значимых и незначимых стимулов 1:1, (Рис.11).

Рис.11 Графики мощности ЗВП при усреднении значимых и незначимых стимулов 1:1. Та же запись, тот же испытуемый. Повышение уровня пиков в области 400-700 мсек.
Если применять усреднение только по значимым стимулам, график мощности имеет максимумы в области 300-500 мсек. Следует обратить внимание, что в данном эксперименте усреднение производилось лишь по 16 стимулам. Тем не менее, график мощности позволяет обнаруживать ожидаемые эффекты (Рис.12).
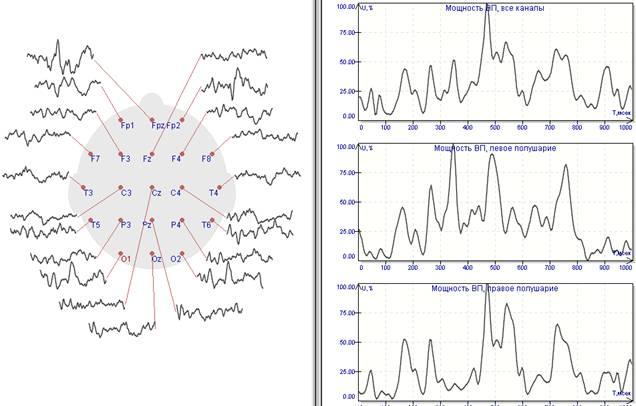
Рис.12 График мощности ЗВП для значимых стимулов. Доминируют пики в областях после 300 мсек., отмечается выраженная межполушарная асимметрия.
Для слуховых вызванных потенциалов можно наблюдать сходные закономерности (Рис.13).

Рис.13 Слуховые вызванные потенциалы (100 усреднений), соотношение значимых и незначимых стимулов – 1:4. На графиках мощности хорошо видна межполушарная асимметрия в областях 350-550 мсек.
Если уменьшить вклад незначимых стимулов при усреднении ВП до 1:1, то компоненты в области 350-550 мсек. проявляются более наглядно (Рис.14)

Рис.14 Усреднение при вкладе значимых и незначимых стимулов в соотношении 1:1. Тот же испытуемый, та же запись. Возрастание уровня поздних компонентов, увеличение асимметрии.
Сопоставление графиков мощности для ЗВП на изменение изображения и слуховых ВП дает возможность наглядно показать отличие функций правого и левого полушарий при обработке акустических сигналов и анализе изображений (Рис.15).


Рис.15 Графики мощности слуховых ВП (слева) и ВП на изменение изображения (справа). Усреднение при соотношении значимых и незначимых стимулов 1:1, при стимуляции соотношение было 1:4 в обоих случаях.
При сопоставлении графиков мощности хорошо видно, что обработка (распознавание) стимула идет преимущественно в структурах левого полушария для слуховых ВП. В то же время распознавание изображений как более сложная задача требует, вероятно, сочетанной работы структур обоих полушарий, хотя асимметрия (связанная, надо полагать, с разделением функций) также присутствует.
Выводы:
1. Вследствие улучшения коэффициента сигнал/шум график мощности ВП дает возможность более точного измерения интервалов времени между откликами.
2. Падение пространственного разрешения не является дефектом метода и может быть при необходимости скомпенсировано традиционным анализом ВП .
3. Метод может быть расширен вычислением дополнительных параметров – например взаимнокорреляционной функции графиков мощности обоих полушарий. В результате могут быть получены не только достоверные данные о корреляции процессов, но и данные об интегральных характеристиках задержки проведения ВП, что принципиально невозможно при существующих методах их анализа.
4. Высокая специфичность графиков мощности для ВП на различные стимулы позволит распознавать тип патологического процесса при проведении комбинированного исследования вызванных потенциалов с разнотипной стимуляцией.
5. Следует ожидать хорошей стабильности формы графика мощности у одного и того же пациента в разные моменты времени исследования.
6. Можно предполагать высокую корреляции вида графика мощности ВП с генетически детерминированными особенностями проведения нервных импульсов (в первую очередь при генетически обусловленных эпилептических синдромах).
7. Нельзя исключить возможность обнаружения патогномоничных форм графика мощности ВП при некоторых нозологиях .
8. Вычисление графика мощности вызванных потенциалов может предоставить новые возможности для анализа функционирования мозговых структур и практической диагностики в условиях клиники.
9. Перечисленные выводы не являются исчерпывающими, т.к. экспериментальная работа находится в начальной фазе и не исключено обнаружение неожиданных феноменов.
VII
Математический анализ биопотенциалов с помощью ЭВМ находит все большее практическое применение. Считается, что без использования ЭВМ нельзя решать сложные задачи, касающиеся расшифровки механизмов кодирования и декодирования информации в головном мозге.
Общепризнанно, что ЭВМ незаменимы тогда, когда требуется сопоставить по времени и по активности значительное количество одновременно протекающих процессов.
Математические методы анализа ЭЭГ являются наиболее результативными и объективными. Их значение особенно возрастает в связи с возможностью использования электронных вычислительных машин, способных быстро выполнять множество громоздких и трудоемких вычислений, что ранее было препятствием широкому применению методов математики для анализа физиологических кривых. Ритмический характер многих процессов, протекающих в живом организме, в определенной степени оправдывает гипотезу о том, что ЭЭГ является результатом алгебраического сложения многих регулярных (например, периодических, синусоидальных и т. п.) колебаний на фоне случайных помех.
В разное время с различным успехом в основном применялись три математических метода для анализа энцефалограмм:
-
гармонический (с помощью рядов Фурье);
-
периодограммный;
-
корреляционный (авто- и кросскорреляционный).
Анализ ЭЭГ с помощью рядов Фурье дает возможность выявить суммарную активность до или после какого-либо раздражения, так как ряд Фурье выделяет гармонические составляющие ЭЭГ с дискретным спектром частот различной амплитуды.
Периодограммный анализ позволяет выявлять скрытые периодичности, т. е. распознавать спектральную структуру естественных процессов по результатам их регистрации. В отличие от других методов анализа ЭЭГ периодограммный метод свободен от таких недостатков, как невозможность учета фаз колебаний, ограничения при анализе быстропротекающих изменений ЭЭГ, наличие артефактов на низких частотах и др. Периодограммный анализ может быть использован также для оценки изменений ЭЭГ под действием различных афферентных раздражителей, а также при фармакологических пробах и др.
Корреляционный анализ дает возможность судить о том процессы каких типов содержатся в данной ЭЭГ, оценит среднюю величину значений периода повторений процесса, степени устойчивости периодического процессах .
Метод
спектрального сводится к вычислению
авто- и кросскорреляционных функции
двух ЭЭГ, одновременно отводимых от
разных точек коры. Кросскорреляционная
функция
![]() считается
так:
считается
так:
 (2)
(2)
В
этой формуле
![]() -
значения двух ЭЭГ в дискретные моменты
времени, отстоящие на интервале
-
значения двух ЭЭГ в дискретные моменты
времени, отстоящие на интервале
![]() и
и
![]() от начала исследуемого отрезка записи;
N –
число интервалов
от начала исследуемого отрезка записи;
N –
число интервалов
![]() на исследуемом отрезке записи;
на исследуемом отрезке записи;
![]() -
интервал квантования; Целочисленная
величина
-
интервал квантования; Целочисленная
величина
![]() может принимать положительные и
отрицательные значения:
может принимать положительные и
отрицательные значения:


Автокорреляционные
функции
![]() ЭЭГ получаются по формуле (2) подставив
значение x
и у
соответственно; в этом случае
ЭЭГ получаются по формуле (2) подставив
значение x
и у
соответственно; в этом случае
![]() принимает только положительные значения.
принимает только положительные значения.
Расчет
спектров мощности автокорреляционной
функции
![]() (автоспектр) и кросскорреляционной
(автоспектр) и кросскорреляционной
![]() (кросс-спектр), а также фазового спектра
(кросс-спектр), а также фазового спектра
![]() производится по следующим формулам:
производится по следующим формулам:
![]()
![]()
![]()
где
![]()
![]()
![]()
где
![]() -
число интервалов
-
число интервалов
![]() в одной ветви кросскоррелограммы; а
в одной ветви кросскоррелограммы; а
![]() -
сглаживающая функция Хемминга
-
сглаживающая функция Хемминга
![]()
в качестве сглаживающей функции можно использовать также функцию Парзена.
Автокорреляционная
функция воспроизводит ритмы, возникающие
в различных участках ЭЭГ, даже если их
фазы в разных участках ЭЭГ произвольно
сдвигаются друг относительно друга.
Это позволяет их анализировать с помощью
преобразования Фурье
![]() .
Кросскорреляционная функция воспроизводит
ритмы одинаковой частоты, появляющиеся
в одних и тех же участках записи в обеих
ЭЭГ, и относительная выраженность этой
ритмики обуславливает кросс-спектр
.
Кросскорреляционная функция воспроизводит
ритмы одинаковой частоты, появляющиеся
в одних и тех же участках записи в обеих
ЭЭГ, и относительная выраженность этой
ритмики обуславливает кросс-спектр
![]() .
Взаимные фазовые сдвиги этих ритмов в
двух ЭЭГ могут быть определены по
фазовому спектру
.
Взаимные фазовые сдвиги этих ритмов в
двух ЭЭГ могут быть определены по
фазовому спектру
![]() [2].
[2].
В зависимости от вида кросскорреляционной функции можно выделить периодические ее оставляющие, общие для двух фиксированных ЭЭГ даже в том случае, если их амплитуды намного меньше амплитуд имеющихся непериодических элементов. Кроме того, можно определить степень связи между амплитудами различных процессов при данном сдвиге времен а также выделить из фоновой активности вызванные потенциалы.
Автокорреляционный анализ используется для изучения степени связи между амплитудами одного и того процесса при данном сдвиге времени.
Анализ спектра мощности методом преобразования Фурье позволяет не только быстро и объективно рассчитать индексы ритмов в выбранных участках записи, но и выявить не заметные на глаз изменения ЭЭГ активности .
Также проводят анализ разности двух ВП. Разность ВП позволяет выяснить меру изменения ВП во времени, что представляет интерес при тестировании влияния различных факторов на ВП (фармакологического воздействия, гипервентиляции и др.). Вычисление разности вызванных потенциалов позволяет получить количественную характеристику различий ВП разных отделов мозга, что важно, например, при выяснении локализации паралогического процесса или при оценке межполушарной функциональной специализации. Асимметрию ВП в гомологических точках разных полушарий легко оценить, используя визуализацию разностного сигнала ВП между какими-либо отведениями .
Для количественной оценке ВП вычисляют площади, ограниченной нулевой линии и кривой ВП в заданном произвольно интервале времени .
Использование методов топографического картирования и трехмерной локализации источников электрической активности позволяет проследить динамику генерации ЭЭГ активности и уточнить локализацию патологического процесса в структурах мозга.
Проследить динамику изменения ВП одного и того же испытуемого или сопоставить ВП разных испытуемых можно с помощью режима по парного сравнения. Такой анализ позволяет сопровождать реабилитационный период, оценить эффективность медикаментозного лечения, сравнить ВП данного пациента с заранее зафиксированной нормой .