

Молекулярная биология гена / В любой соматической клетке человека 23 пары хромосом
.doc-
В любой соматической клетке человека 23 пары хромосом. В каждой из них по одной молекуле ДНК. Длина всех 46-и молекул почти 2 м.
-
Сколько же всего генов, то есть последовательностей нуклеотидов, кодирующих белки, в ДНК человека?
По последним данным – генов у человека примерно 25 тыс. На их долю приходится только 2% от общей ДНК. Остальная ДНК – некодирующая – «мусорная».
-
Вставив в ДНК мыши «мусорный» фрагмент человеческой ДНК, не кодирующий ни одного белка, можно сделать её лапку похожей на человеческую кисть.
-
Обнаружено, что почти все человеческие гены кодируют более одного белка
-
Альтернативный сплайсинг — механизм редактирования молекул РНК, благодаря которому на основе одного и того же гена организм может синтезировать несколько вариантов (изоформ) белковой молекулы.
-
Анализ 400 млн фрагментов РНК из разных тканей и органов показал, что 94% человеческих генов подвергаются альтернативному сплайсингу, причем в разных тканях производятся разные наборы изоформ.
Благодаря альтернативному сплайсингу разнообразие белков в организме млекопитающих значительно выше, чем у низших животных, хотя количество генов у тех и других примерно одинаково.
В нашем геноме оказалось около 25 тыс генов — примерно столько же, сколько у круглого червя Caenorhabditis elegans, который устроен гораздо проще, чем человек.
-
Во время удвоения ДНК могут возникать различные ошибки: не только замена одного нуклеотида на другой, но и перенос, удвоение или потеря участка ДНК. Большинство таких изменений не затрагивает жизненно важные гены и поэтому не отсеивается естественным отбором. В результате в любой крупной популяции (например, человеческой) обнаруживается много структурных вариантов генома (СВГ).
Отличие ДНК от РНК
• В ДНК при втором углеродном атоме (C2') находится атом водорода (Н), в РНК – гидроксильная группа (ОН). Эта группа увеличивает вероятность гидролиза молекулы РНК, то есть уменьшает ее стабильность.
В виду того, что в С2' положении РНК находится ОН-группа, двухнитчатая структура РНК образует А-форму двойной спирали.
• С2'-ОН группа делает РНК не только более конформационно пластичной, но также обеспечивает способность образовывать дополнительные водородные связи (что дает трехнитчатые участки) и взаимодействовать не только с фосфорной кислотой, но и с ионами металлов.
Биологический смысл репликации ДНК: копирование генетической информации для переноса ее следующей генерации
-
Репликация происходит с помощью полуконсервативного синтеза: двойная спираль раскручивается,каждая родительская цепь служит в качестве матрицы для синтеза дочерней цепи,в ходе синтеза дочерних цепей возникают новые комплементарные пары, в результате репликации образуются две новые одинаковые дочерние цепи.
-
Ферменты репликации
-
Для подготовки хромосомы к репликации - топоизомеразы
Ферменты, непосредственно участвующие в процессе репликации: Хеликазы;Белки инициации репликации DnaА,DnaB,DnaС;Ssb-белки;ДНК-праймаза;ДНК-полимеразы:(ДНК-полимераза I, ДНК-полимераза II, ДНК-полимераза III);ДНК-лигаза
-
E. coli имеет три ДНК-полимеразы: ДНК полимераза I (pol I),ДНК полимераза II (pol II),ДНК полимераза III (pol III) - основной фермент репликации ДНК. Все три полимеразы обладают полимеризующей активностью в направлении 5 ′ 3′. Кроме того, все три полимеразы имеют 3′ 5′ экзонуклеазную активность, которая устраняет собственные ошибки при репликации. Фермент ДНК pol I имеет дополнительно экзонуклеазную активность для удаления РНК-праймера в направлении 5 ′ 3′.
Репликация у эукариотических клеток осуществляется также по полуконсервативному типу, однако имеет свои особенности:
-
Репликация по типу множественных глазков при репликации хромосом эукариотических организмов.
-
Более сложная система ферментов для репликации ДНК хромосом и наличие проблемы укорочения теломерных концов.


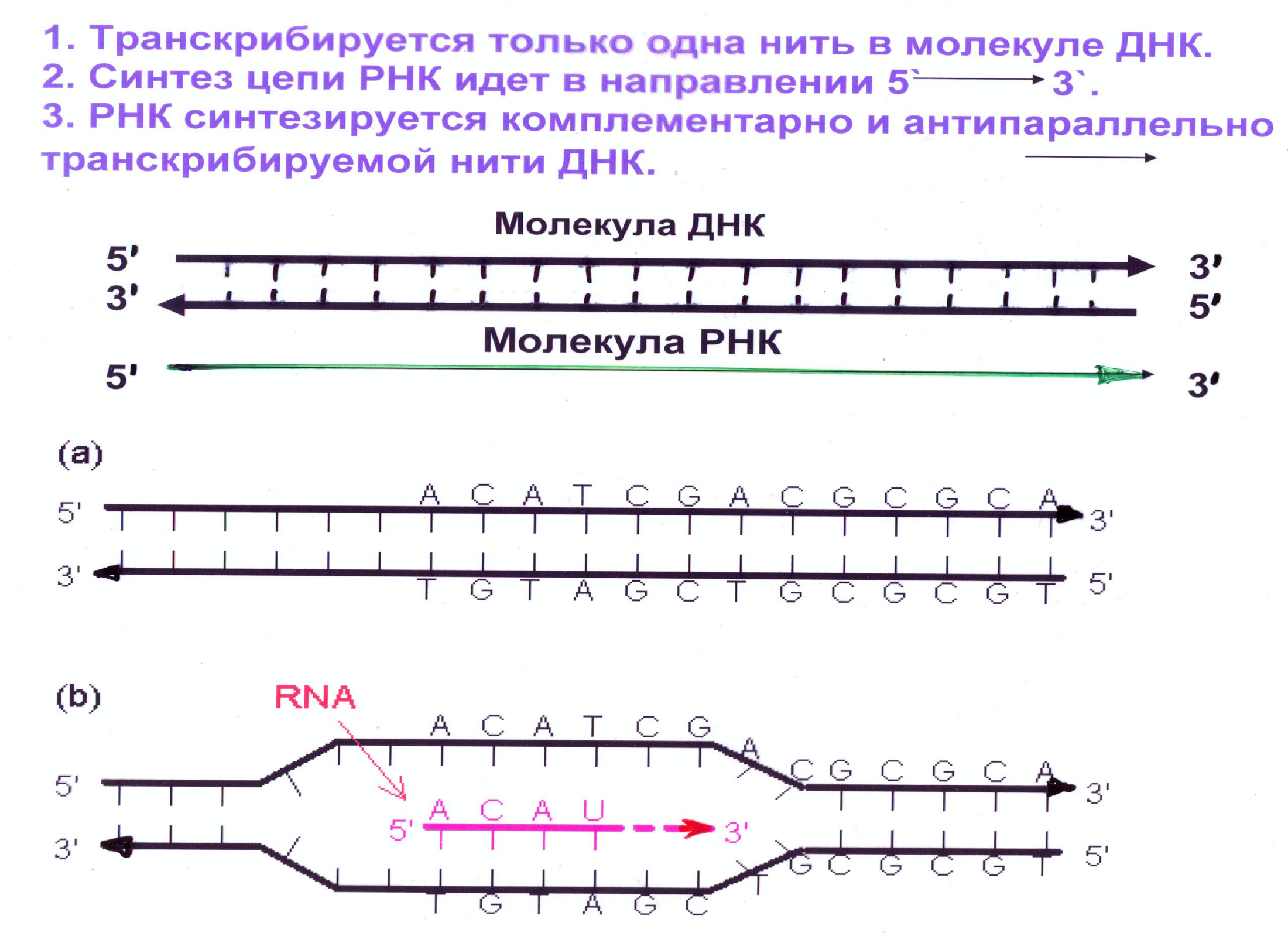
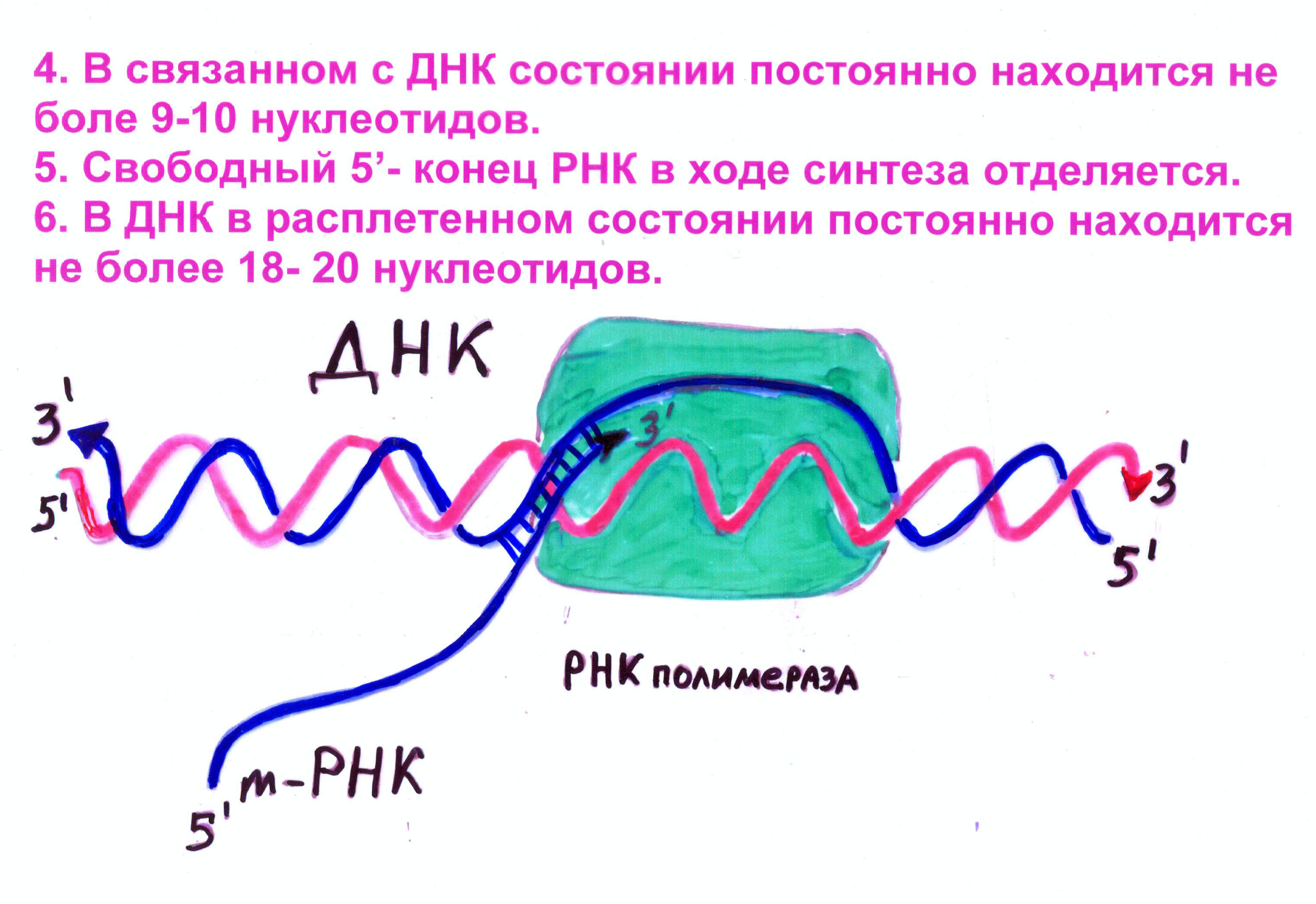



Четыре типа РНК-полимераз у эукариот (3 типа ядерные и 1 тип – митохондриальная или хлоропластная)
-
РНК Pol I - Синтезирует три типа рРНК (28S, 18S, 5,8S);
-
РНК Pol II – синтезирует мРНК и малые ядерные РНК;
-
РНК Pol III – синтезирует тРНК и 5S-РНК.
РНК-полимеразы
эукариот имеют большую молекулярную
массу и представляют собой комплекс
мультимерных белков (500 – 700 кД). От 14 до
17 субъединиц в зависимости от типа
полимера Прокариотические
промоторы варьируют в размерах – от 20
до 200 п.н., но наиболее типичным является
промотор, величиной 40 п.н. Внутри промотора
имеется две постоянные последовательности
(консенсусные последовательности). Эти
элементы называются Прибнов-боксом или
«-10-м боксом» и «-35 боксом».Эукариотические мРНК
довольно стабильны.
Период их полураспада измеряется часами
и даже сутками. Эукариотические
мРНК синтезируются в виде предшественников
и проходят в своем биогенезе стадию
довольно сложного созревания, или
процессинга.
Прокариотические
промоторы варьируют в размерах – от 20
до 200 п.н., но наиболее типичным является
промотор, величиной 40 п.н. Внутри промотора
имеется две постоянные последовательности
(консенсусные последовательности). Эти
элементы называются Прибнов-боксом или
«-10-м боксом» и «-35 боксом».Эукариотические мРНК
довольно стабильны.
Период их полураспада измеряется часами
и даже сутками. Эукариотические
мРНК синтезируются в виде предшественников
и проходят в своем биогенезе стадию
довольно сложного созревания, или
процессинга.
Процессинг включает в себя:
-
Кэпирование 5'-конца, заключающееся в присоединении к этому концу мРНК так называемой шапочки (кэп-структуры – 7-метил-гуанозина),
-
Полиаденилирование 3'-конца (образование polyA-хвоста, длиной до 200 нуклеотидов.
3. Сплайсинг - вырезание протяженных внутренних участков мРНК, так называемых интронов, и ковалентное воссоединение оставшихся фрагментов (экзонов) через обычную фосфодиэфирную связь.
Механизм транскрипции:
-
Узнавание промотора
-
Инициация
-
Элонгация
-
Терминация
Самые важные составляющие транскрипционного комплекса: РНК-полимераза и пять белков-помощников (главные факторы транскрипции – TFIIB, D, E, F и H-факторы), без которых РНК-полимераза не способна узнать промотор и начать транскрипцию.В транскрипции эукариот принимает участие также белковый комплекс, названный Медиатором. Его предназначение – определять, какие именно белки будет строить клетка. Медиатор передаёт полимеразе сигнал от энхансера, который может воспринимать сигналы о том, какие белки нужны клетке в данный момент и усиливать работу гена или группы генов. В инициации транскрипции, осуществляемой РНК-полимеразой II, участвуют более 60 белков – транскрипционных факторов.
Гриб бледная поганка (Amanita),содержит яд α-аманитин, блокирующий работу РНК-полимеразы II. Этот фермент очень чувствителен к α-аманитину (концентрация 10 М полностью подавляет работу фермента). Несколько меньшая чувствительность у РНК-полимеразы III (работа фермента подавляется в концентрации яда 10 М), РНК-полимераза I – нечувствительна к α-аманитину.

Рифампицин прикрепляется к β-субъединице РНК-полимеразы и блокирует синтез РНК. Этот антибиотик не запрещает образование новой фосфодиэфирной связи, но блокирует транслокацию РНК-полимеразы вдоль ДНК-матрицы.
Трансляция – это осуществляемый рибосомами синтез белка из аминокислот на матрице мРНК (или иРНК).
Cоставляющие элементы процесса трансляции: аминокислоты, тРНК, рибосомы, мРНК, ферменты для аминоацилирования тРНК, белковые факторы трансляции (белковые факторы инициации, элонгации, терминации - специфические внерибосомные белки, необходимые для процессов трансляции), источники энергии АТФ и ГТФ, ионы магния (стабилизируют структуру рибосом).
Размер тРНК примерно 80 нуклеотидов. тРНК действуют как адапторы между кодонами на мРНК и аминокислотами, которые они кодируют.
Для каждой из 20 аминокислот в клетках есть фермент, осуществляющий синтез соответствующей аминоацил-тРНК (общее название - аминоацил-тРНК-синтетаза).
-
Каждая тРНК специфична к одной аминокислоте и к одному кодону мРНК
-
Аминокислота активируется (с использованием АТФ) и присоединяется к тРНК с помощью фермента аминоацил-тРНК синтетазы
-
АК + тРНК + АТФ = АК-тРНК + АМФ + ФФ
-
включающую две стадии:
-
Фермент + АК + АТФ = Комплекс (фермент(АК-АМФ) + ФФ (1)
-
Комплекс (фермент(АК-АМФ) + тРНК = фермент + АМФ + АК-тРНК (2)
Чтобы
аминокислота «узнала» свое место в
будущей полипепетидной цепи, она должна
связаться с транспортной
РНК (тРНК),
выполняющей адапторную функцию. Затем
тРНК, связавшаяся с аминокислотой
«унает»соответствующий кодон на мРНК.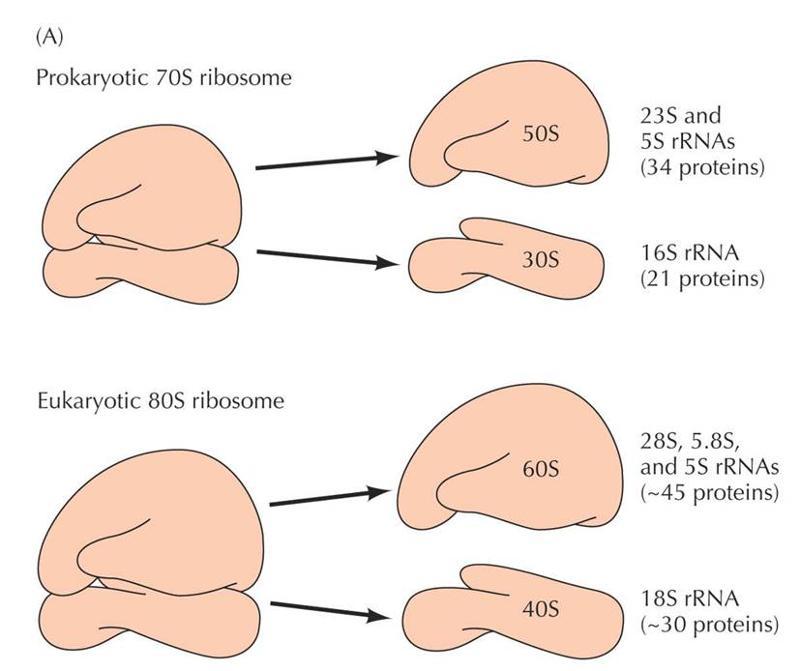
-
Р – пептидильный участок для пептидил-тРНК
-
А – аминоацильный участок для аминоацил-тРНК
-
Е – участок для выхода тРНК из рибосомы
На стадии инициации в процессе трансляции у бактерий E.coli участвуют три белковых фактора – IF-1, IF-2 и IF-3.IF-3-фактор прикрепляется к 30S-субчастице рибосомы и способствует его взаимодействию с мРНК. F-1-фактор закрывает А-сайт на 30S-субчастице рибосомы, обеспечивая тем самым посадку первой fMet-tRNAfMet на Р-сайт рибосомы и защищая А-сайт от посадки какой-либо другой нагруженой аминокислотой TРНК.IF-2 – это маленький ГТФ связывающий белок, который в форме IF-2-ГТФ прикрепляется к нагруженой метионином fMet-tRNAfMet и помогает ей сесть на рибосому (на малую субчастицу).Кроме того, IF-3 помогает tRNAfMet правильно осуществить взаимодействие с инициирующим кодоном AUG на мРНК.Когда большая субчастица рибосомы прикрепляется к малой субчастице, происходи гидролиз ГТФ, что приводит к диссоциации IF2-ГДФ и диссоциации IF1. Таким образом, большая субчастица рибосомы служит как «ГТФ-аза активирующий белок» для IF-2.Когда две субчастицы рибосомы оказываются соединенными вместе, мРНК оказывается в канале, который образуется между субчастицами рибосомы.Второй этап синтеза белка – элонгация, требует участия факторов элонгации: EF-Tu,EF-Ts,EF-G. EF-Tu и EF-G являются маленькими ГТФ-связывающими белками.
Генетический код – это единая система записи наследственной информации в виде последовательности нуклеотидов в ДНК или мРНК, которая определяет последовательность аминокислот в белке
Свойства генетического кода:
1. Триплетность: каждая аминокислота кодируется триплетом нуклеотидов в ДНК или мРНК.
2. Однозначность: один кодон соответствует одной аминокислоте.
3. Непрерывность: кодоны мРНК не отделены друг от друга (отсутствуют «запятые»).
4. Вырожденность (избыточность): одна аминокислота может кодироваться разными кодонами.
5. Неперекрываемость: каждый нуклеотид в мРНК принадлежит только одному кодону (исключения обнаружены у вирусов).
6. Универсальность: генетический код одинаков для всех организмов.
7. Колинеарность: последовательность триплетов в ДНК соответствует последовательности аминокислот в белке.
8. Инициирующим кодоном является АУГ (редко ГУГ).
9. Терминирующими кодонами являются УАА, УГА, УАГ.
Генетический код триплетный: каждая аминокислота кодируется триплетом нуклеотидов в ДНК и соответствующим кодоном мРНК. Первое предположение о строении генетического кода появилась уже в 1954 г. Георгий Гамов предположил, что кодирование информации в молекулах ДНК осуществляется сочетаниями нескольких нуклеотидов. Очевидно, что однозначного соответствия «1 нуклеотид - 1 аминокислота» в коде быть не может, так как в этом случае должно было быть только 4 аминокислоты . Дуплетный код «2 нуклеотида – одна аминокислота» соответствовал бы только 16-и аминокислотам. Лишь триплетный код обеспечивает шифровку 64 аминокислот (т.е. 20 аминокислот могут кодироваться триплетным кодом).
В 1957 году Ф.Крик и С. Бреннер экспериментально доказали, что генетический код триплетен. Эксперименты проводили с бактериофагом Т4 и бактериями E.coli. У бактериофага Т4 индуцировали мутации со сдвигом рамки считывания в локусе rII с помощью акридиновых красителей. Мутации сдвига рамки считывания показали, что код триплетный.
Генетический код однозначный: один кодон соответствует одной аминокислоте
В 1961 г. биохимики М. Ниренберг и Г. Маттеи изучали синтез белков в бесклеточной системе E.coli. В каждой из 20 пробирок имелись все клеточные компоненты бактерий (кроме нуклеиновых кислот) и все 20 аминокислот, одна из которых содержала радиоактивную метку. В одном из экспериментов в качестве матрицы добавили в реакционную смесь полиуридиловую кислоту (РНК, состоящую из UUUUUUU). В результате в 1000 раз увеличилось количество белка, содержащего меченую аминокислоту фенилаланин. Был сделан вывод, что фенилаланин кодируется трилетом UUU. Подобным образом было показано, что кодон ААА кодирует лизин, кодон ГГГ – глицин.
В 1964 г М. Ниренберг и Ф. Ледер разработали метод связывания на рибосомах в бесклеточной системе аминоацил-т-РНК с мРНК матрицами, состоящими из 2-х и 3-х нуклеотидов) с заданным порядком оснований.
Все динуклеотиды оказались неэффективными в связывании т-РНК. Тринуклеотиды же обеспечивали присоединение аминоацил-т-РНК к матрице РНК. Тем самым окончательно была доказана триплетность кода.
Полученные комплексы подвергали фильтрованию через мелкопористый фильтр, пропускавший отдельные компоненты, связавшиеся аминоацил-т-РНК, нагруженные определенной аминокислотой оседали на фильтре. Эти комплексы мини-т-РНК + аминокислота позволили определить соответствие между аминокислотой и типом мини-т-РНК. Метод позволял перепробовать и все возможные триплеты по одному. В результате уже через год были испытаны все 64 триплета. Почти все они были признаны кодирующими. Было показано, что из 64 возможных триплетов 61 кодирует различные аминокислоты; а 3 триплета (УАА, УГА, УАГ) получили название бессмысленных, или «нонсенс-триплетов» Они не кодируют аминокислот, но выполняют функцию знаков препинания при считывании наследственной информации.
Генетический код непрерывный: кодоны на мРНК не отделены друг от друга (отсутствуют «запятые»)
Важнейшими характеристиками генетического кода являются его непрерывность и неперекрываемость кодонов при считывании. Это означает, что последовательность нуклеотидов считывается триплет за триплетом без пропусков, при этом соседние триплеты не перекрывают друг друга, т.е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания.Доказательством неперекрываемости генетического кода является замена только одной аминокислоты в пептиде при замене одного нуклеотида в ДНК. В случае включения нуклеотида в несколько перекрывающихся триплетов его замена влекла бы за собой замену 2 или 3-х рядом стоящих аминокислот в пептидной цепи.
Генетический код вырожденный (избыточный): одна аминокислота может кодироваться несколькими кодонами
Обращает на себя внимание избыточность кода, проявляющаяся в том, что многие аминокислоты шифруются несколькими триплетами. Это свойство триплетного кода, названное вырожденностью, имеет очень важное значение: мутационные изменения молекулы ДНК типа замены одного нуклеотида на другой далеко не всегда изменяет смысл триплета. Возникшее таким образом новое сочетание из трех нуклеотидов часто кодирует ту же самую аминокислоту. Иными словами, вырожденность кода повышает его запас прочности в случае возникновения генных мутаций.
Генетический код универсальный – он практически одинаков для всех организмов
Универсальность генетического кода свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции.Отличия генетического кода обнаружены у митохондрий и пластид. Это свидетельствует в пользу дивергентности эволюции кода на ранних этапах существования жизни. В митохондриальных ДНК различных видов организмов отмечаются некоторые отклонения от генетического кода.
Колинеарность генетического кода: последовательность триплетов в ДНК соответствует последовательности аминокислот в белке.