

Fiziologia_rasteny
.pdf2.В клетке происходит синтез предшественников лигнина, которые участвуют в разрушении ауксина и торможении клеточ- ного растяжения.
3.В клетке на последнем этапе растяжения синтезируется в большом количестве этилен — антагонист ауксина и ингибитор клеточного растяжения.
Растянутая клетка с большой центральной вакуолью переходит
êследующему этапу жизнедеятельности — дифференцировке. Дифференцировкой называют превращение эмбриональной
клетки в специализированную. После окончания роста растяжением отдельные клетки развиваются разными путями.
Каждое состояние дифференцировки клетки характеризуется определенной генной моделью, вызывающей эту дифференцировку через индукцию специфических ферментов. Дифференцировка — появление качественных различий между клетками, тканями и органами в процессе развития.
Когда дифференцировку клеток изучают по морфологическим признакам, тогда говорят о дифференцировке структурной. Когда разговор идет о формировании в клетках отличий в составе белков-ферментов, в способности к синтезу запасных или других веществ и других биохимических изменениях, дифференцировку называют биохимической.
Дифференцировка клеток приводит к возникновению как специфической формы, так и специализации выполняемых функций. Различают и физиологическую дифференцировку (разница между корнями и побегом, между вегетативными и репродуктивными фазами жизненного цикла).
Как правило, дифференцированные клетки объединены в ткани, т. е. образуют группы клеток, выполняющих определенную физиологическую функцию и имеющих схожее морфологическое строение, обеспечивающее реализацию этой функции.
Существуют разные классификации типов дифференцированных клеток, одну из них можно представить в следующем виде:
паренхимные, характеризуемые большими размерами, тонкими оболочками, содержанием хлоропластов или запасных веществ;
проводящие и поддерживающие — все клетки этой группы вытянуты, часть из них сильно лигнифицирована, представлена трахеидами, сосудами и волокнами. Живое содержимое в них почти отсутствует;
332
покровные — обычно находятся на поверхности и покрыты водонепроницаемыми веществами (воском, кутином, суберином).
Êним относится эпидермис, ризодермис и перидерма;
репродуктивные, образующиеся в определенные периоды жизненного цикла растений, из которых потом формируются гаметы, необходимые для полового размножения высших растений.
Важным вопросом клеточной дифференцировки является вопрос о механизмах, которые лежат в основе этого явления. На- чальным этапом дифференцировки является образование физиологической оси с двумя полюсами. Дальнейшая дифференцировка многоклеточного организма определяется дифференцированной реализацией генетической информации во времени и пространстве, содержащейся в генотипе клетки.
Таким образом, в индукции дифференцировки первыми шагами служит возникновение полярности. Полярность индуцируется градиентом какого-либо фактора окружающей среды. Фактор может иметь физическую (свет, сила тяжести, электриче-
ское поле, температура) или химическую (фитогормоны, ионы Са2 ) природу.
Возникшая полярная ось — необходимая предпосылка для поддержания внутриклеточных градиентов.
В многоклеточном организме значительную роль в дифференцировке играет передача информации между клетками. У растений наиболее изучена гормональная передача информации и в меньшей мере электрофизиологический способ передачи информации. Начав дифференцироваться, клетки не только изменяются по структуре, но и занимают определенное место в ассоциации себе подобных, образуют протканевую структуру.
Соседство клеток одна с другой обеспечивает программу дифференцировки и рост клеточной ассоциации. Контакты растущих клеток в протканевых фрагментах происходят не только за счет поверхностных агентов, но и при участии внутриклеточных компонентов. Большую роль при этом играют микротрубочки, состоящие из подобного актину белка — тубулина.
Сформировав протканевую структуру, клетки начинают свое кооперативное движение: пласт клеток перемещается один относительно другого, образуя первичную ткань.
Дифференцировка клеток во вновь образованной ткани происходит в два этапа. Сначала формируется одна из специализиро-
333
ванных клеток, затем возникают ей подобные. Большую роль в процессе тканевой дифференцировки играют фитогормоны.
В растущем органе, каким является, например, лист, формирование тканей происходит неодновременно. Прежде всего клеточное деление заканчивается в эпителиальной и проводящей тканях, затем наступает процесс активного клеточного растяжения и дифференцировки. После этого подобные процессы происходят в мезофилле листа.
Формирование органа, таким образом, происходит за счет последовательной дифференцировки отдельных тканей. Однако конечный размер органа — комплексный результат роста его отдельных тканей и клеток, т. е. размер и форма органа предопределяются в меристеме.
Будущая дифференцировка зависит от того, в какой части меристемы инициальная клетка находится. Когда меристематиче- ская клетка локализована в органогенной зоне, то из нее образуется группа клеток, составляющая лист, клетки субапикальной зоны формируют стебель и т. д.
Следовательно, уже в меристематической зоне происходит своеобразный процесс детерминации, в результате которого клеточная система выбирает один из многих путей развития.
Если коротко отметить, то дифференцировка растительных клеток включает в себя индукцию полярности и дифференциальной активности генов, в результате клетка детерминируется и приобретает черты специализации. В качестве индукторов дифференцировки, как отмечалось, выступают как физические, так и химические факторы внутренней и внешней среды. Причем каждая клетка непрерывно получает информацию о своем окружении и развивается в соответствии с этой информацией.
Детерминация — это определение пути дифференцировки клетки. При детерминации делается выбор из большого количе- ства потенций (генов, информации) в определенном направлении. Детерминация клетки может быть запрограммирована или возникает под воздействием разных внешних факторов: соседних клеток, гормонов и т. д.
Важную роль в детерминации будущей дифференцировки играет клеточное окружение. Пересадка одной клетки из группы эмбриональных клеток в область со специфическими функциями может полностью изменить будущую программу развития этих клеток. Особенно хорошо эти опыты удаются с зародышами насе-
334
комых. Так, клетки будущего глаза превращаются в клетки крыла насекомых и т. д.
Растениям принадлежат интенсивные регенерирующие способности. Черенок в определенных условиях способен давать целое растение, но такой же регенерирующей способностью обладает лист (листовой черешок) и, наконец, часть клетки — протопласт. Проходя через ряд промежуточных фаз, протопласты становятся клетками, регенерируя оболочку.
Это связано с уникальной способностью растительной клетки — под влиянием воздействий реализовать принадлежащую ей тотипотентность и дать начало целому организму. Тотипотентной является любая клетка растения, так как она владеет полным генофондом, т. е. всеми возможностями будущего организма. Тотипотентные клетки — генетически однородные клетки.
Все органы растительного организма взаимосвязаны и влияют на рост друг друга. Влияние одних частей организма на скорость и характер роста других, часто на большие расстояния, называют корреляцией. Корреляция обусловливает упорядоченную взаимозависимость отдельных частей растения. Корреляции можно сравнить с отношениями между клетками, перенесенными на уровень тканей и органов.
Включая в себя дальний транспорт, корреляции связаны с действием гормонов (хотя не каждая корреляция имеет гормональный характер). Простейший пример корреляции — ускорение роста в зоне растяжения колеоптиля ауксином, поступающего из его верхушки.
Только в редких случаях один гормон имеет решающее значе- ние для корреляции, а чаще всего необходимо количественное соотношение нескольких гормонов. В полярных, однонаправленных воздействиях почти всегда участвует полярно транспортируемый ауксин.
Имеет место как коррелятивная стимуляция, так и коррелятивное торможение. В первом случае растение с более мощной корневой системой благодаря большему поступлению питательных веществ имеет и лучший рост побегов; побег влияет на корень, поставляя ему ауксин, а корень действует на побег при помощи цитокининов и гиббереллинов. Во втором случае размер плодов уменьшается с увеличением их количества; апикальное доминирование — верхушечный побег тормозит развитие боковых; удаление верхушечного побега приводит к развитию боковой почки, т. е. происходит разветвление стебля.
335
6.4. Онтогенез и его этапы
Рост растений — сложный процесс, в его основе лежат такие фундаментальные явления, как ритмичность, полярность, дифференциация, раздражимость, корреляция. Все эти процессы общие для онтогенеза всех живых организмов.
Онтогенез — индивидуальное развитие организма от зиготы (или вегетативного зачатка) до природной смерти. Включает в себя все жизненные процессы и продолжается у разных растений от 10—14 дней до 3—5 тыс. лет. Самыми долгоживущими являются секвои — до 5 тыс. лет.
В ходе онтогенеза рост и развитие взаимосвязаны и локализованы в одних и тех же тканях — меристемах.
По продолжительности жизни растения делят на эфемеры, однолетние, двулетние и многолетние.
Эфемеры — растения, очень быстро проходящие свое развитие от прорастания до плодоношения. Особенно много эфемеров в полупустынях и пустынях.
Однолетние — это растения, живущие одно лето или один год;
двулетние — äâà ãîäà; многолетние — много лет.
Независимо от продолжительности жизни все растения подразделяют на две группы: моно- и поликарпические. Монокарпи- ческие цветут и плодоносят один раз в своей жизни; поликарпи- ческие — много раз в жизни.
Благодаря активной деятельности меристем и фотосинтетиче- ской активности листьев зеленое растение приобретает ряд тех своеобразных черт, которые характеризуют его рост. Последний в процессе онтогенеза растения наблюдается на протяжении основных этапов его жизненного развития (цикла).
Этапы онтогенеза
Этапность онтогенеза — морфологическое и функциональное разветвление онтогенеза, проявляется в изменении характера роста, дифференциации и функциональной активности организма. Таких этапов выделяют пять: эмбриональный, ювенильный, спелость, размножение и старость.
Этапы онтогенеза — не изолированные периоды развития, а взаимопереходящие одна в другую фазы, в основе которых лежат медленно идущие возрастные изменения, т. е. изменения организма и его отдельных частей, обусловленные возрастом и идущие на протяжении всей жизни растений. Возрастные изменения
336
клеток, тканей и органов зависят от присущего им роста, общего возраста организма, от характера взаимосвязей с другими частями растения.
Эмбриональный этап у растений, размножающихся семенами, — это период формирования зародыша и семени от оплодотворения яйцеклетки до начала прорастания семени; для вегетативно размножающихся растений — период формирования почек
âорганах вегетативного размножения от возникновения почки до начала ее прорастания. В это время ростовые процессы находятся
âскрытой фазе или фазе подготовки. Происходит синтез основных метаболитов, ядра, образование ростовых гормонов.
Ювенильный этап — период заложения, роста и развития вегетативных органов от прорастания семени до появления способности к образованию репродуктивных органов. В начале ювенильного периода растения не переходят к образованию репродуктивных органов даже в оптимальных условиях; потом они постепенно приобретают способность к репродукции, т. е. это этап доминирования ростовых процессов. В это же время растение и его органы активно увеличиваются в размерах, происходит новообразование отдельных элементов растения (клеток, тканей, органов). Многие авторы отмечают взаимодействие фитогормонов в период интенсивного роста растительного организма.
Спелость (зрелость) — период цветения в семенных или репродукции в вегетативно размножающихся растениях, от появления первичных зачатков репродуктивных органов до формирования бутонов, цветков, клубней, луковиц и других органов, образование новых зародышей. В этот период процесс роста сопряжен с элементами генеративного развития. Совместно с ростом вегетативных частей растения происходит детерминация генеративных органов — приобретение клеткой, тканью, органом или организмом состояния готовности к реализации определенных наследственных свойств. Детерминация развития характеризуется готовностью к развитию по определенному типу. Другими словами, растение переходит от этапа вегетативного размножения к этапу генеративного, т. е. к этапу спелости.
Размножение — период плодоношения, роста, развития и поспевания плодов и семян в растениях, размножающихся семенами; или поспевание клубней, луковиц и других органов у вегетативно размножающихся растений. Во время этого этапа доминируют процессы роста генеративных органов, увеличение размеров семян или других органов. В этот период совместно с гормонами выявляются и доминируют природные ингибиторы.
337
Старость — последний этап — период от полного прекращения плодоношения до природного отмирания растений. Рост в этом слу- чае происходит очень редко (поросль из глубоко спящих почек, жирующие побеги). Замедляют старение факторы, стимулирующие синтез РНК и белков, и в первую очередь это цитокинины.
Отметим, что в онтогенезе наблюдаются этапы как интенсивного роста, так и его замедление (покой).
6.5. Гормональная регуляция роста
Кроме поступаемых из почвы воды и минеральных веществ, углеводов, образующихся в процессе фотосинтеза, необходимых в качестве источника энергии и строительных белков протоплазмы, растительная клетка для оптимального роста требует еще и некоторые другие химические соединения. К ним, в частности, относятся органические соединения — гормоны. Потребность в количе- стве гормонов обычно очень мала, и в большинстве случаев гормоны синтезируются в достаточных количествах самим растением.
Более того, между клетками в организме должен происходить обмен информацией. Одна группа клеток «отправляет» сигнал, другая — воспринимает его. Молекулу химической природы, обладающую сигнальной функцией, называют первичным мессенджером. Среди обширного спектра первичных мессенджеров выделяют и гормоны растений.
Признаки отнесения вещества к гормонам:
вещество вызывает специфический физиологический ответ; особенность растительных гормонов в том, что они запускают крупные программы развития не только клеток, но и на уровне тканей, органов, целого растения;
синтезируется в растении одной группой клеток, а отвечает за него другая группа (разобщено место синтеза и место действия,
ò.е. сигнальное вещество транспортируется). К синтезу гормонов потенциально способна любая клетка растений; как правило, фитогормоны являются низкомолекулярными соединениями (не более 2 кДа);
практически не играет роли в основном метаболизме клетки, а используется лишь для сигнальных целей.
действует в низкой концентрации.
Первичные мессенджеры, воспринимаемые клеткой, специфически взаимодействуют со многими молекулами-мишенями, в
338
частности с рецепторами. Чтобы клетка ответила на стимул, нужно включить внутриклеточную систему вторичных мессенджеров, усиливающих сигнал в десятки и сотни раз. Например, одна молекула ауксина активирует до 104 протонов.
Критерии обнаружения рецепторов:
высокая избирательность и структурная специфичность в отношении гормона; чем больше физиологический ответ, тем выше аффинность;
эффект проявляется при низких концентрациях гормона;
кинетика связывания с агонистом описывается кривой с насыщением;
при взаимодействии с рецептором не должна меняться химическая структура гормона;
взаимодействие гормона с рецептором приводит к включе- нию системы вторичных посредников.
Большинство систем вторичных мессенджеров, изученных у животных, обнаружены и у растений. Это фосфатидилинозитольная и аденилат- и гуанилатциклазные системы.
Рецепторы обычно локализованы в плазмалемме, но могут находиться и в других компартментах клетки. Рецептор связан с вторичными мессенджерами. Передача сигнала с помощью вторичных посредников активизирует процессы фосфорилирования/дефосфорилирования, что приводит к изменению метаболизма и работы цитоскелета.
Итак, любой гормон представляет собой вещество, образуемое
âмалых количествах в одной части организма и транспортируемое затем в другую часть растения, где он оказывает соответствующее действие. Расстояние, на которое транспортируется гормон, может быть относительно большим, например от корня до листа, от последнего к почке, может быть и меньшим — от апикальной меристемы до находящихся ниже клеток, или совсем малым — в пределах одной клетки.
Основные классы регулирующих рост гормонов
У высших растений содержатся важные классы регулирующих рост гормонов, основные из которых: ауксин, гиббереллины, цитокинины, абсцизовая кислота и этилен.
Ауксин
Â30-õ гг. ХХ столетия Ф. Венту удалось подтвердить мысль
×.Дарвина, что в верхушках колеоптилей злаков действительно образуется значительное количество способного к диффузии ве-
339
щества, которое контролирует рост нижележащих зон. Это вещество назвали ауксином (от греч. auxano — расти). Большой вклад в изучение механизма действия ауксина внес Н. Г. Холодный.
Ауксин синтезируется растущими апикальными зонами стеблей, в том числе молодыми листьями. От апекса ауксин мигрирует в зону растяжения, где он специфически влияет на рост растяжением.
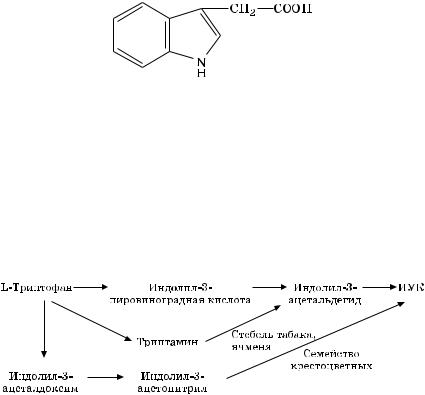
Природный ауксин представляет собой простое соединение — индолил-3-уксусную кислоту (ИУК):
Она синтезируется в растениях путем ферментативного превращения аминокислоты — триптофана.
Триптофан образуется из шикимовой кислоты через ряд реакций:
шикимовая кислота индол серин L-триптофан.
Образованная из триптофана ИУК проходит дальнейшее превращение в растениях: обратимое связывание ИУК в неактивные комплексы с белками, аминокислотами, углеводами; необратимое окисление молекулы ИУК под воздействием ферментов типа ауксиноксидазы. Синтез ИУК из триптофана — сложный процесс, и он имеет свои особенности в отдельных растениях (рис. 6.2).
Ðèñ. 6.2. Биосинтез индолил-3-уксусной кислоты
Транспорт ИУК происходит полярно со скоростью 10—15 см/ч от вершины побегов к корням. Механизм полярного транспорта следующий: в апикальный конец клетки ИУК проникает пассив-
340

но совместно с Н , а в базальном конце — активно секретируется через клеточную мембрану.
Физиологическое действие ауксина сложное. Разные ткани отвечают на действие ауксина увеличением роста, которое обусловлено стимуляцией растяжения клеток (рис. 6.3).
Ðèñ. 6.3. Схема основных функций ауксина в растении
Аттрагирующее действие заключается в том, то клетки и ткани, обогащенные ауксином, становятся центрами притяжения веществ.
Роль ауксина в стимуляции опадания листьев и цветов связана с заметным понижением его содержания в листьях.
Это приводит к старению листьев. Стареющие ткани продуцируют этилен, который действует на отделительную зону (зона опадания).
Сам же ауксин задерживает опадание листа и цветов. Есть доказательства, что ауксин, кроме участия в растяжении клеток и опадании листьев, стимулирует процессы клеточного деления. Вероятно, ауксины повышают камбиальную активность. Считают, что ауксины участвуют в дифференциации сосудистой ткани в период начала ростового процесса и в образовании боковых корней. Превращение завязи в плод — еще один контролируемый ауксином процесс.
Первостепенную роль играет ауксин в ростовых движениях — тропизмах и настиях (теория гормональной регуляции тропизмов Холодного — Вента).
В основе действия ауксинов существует два механизма: быстрое влияние ауксинов на мембранную систему, где за счет энергии АТФ увеличивается транспорт водородных ионов из цитоплазмы в клеточную оболочку и ускоряется размягчение клеточной оболочки; медленное влияние ауксинов через геномную систему на синтез белков, определяющих рост клеток.
Наличие обоих механизмов весьма вероятно, поскольку ауксин не только вызывает выход протонов, но и изменяет микроструктуру цитоплазмы (микротрубочки).
341