

Fiziologia_rasteny
.pdf
|
|
|
Окончание табл. 5.2 |
|
|
|
|
|
|
Элемент |
Концентрация в сухом веществе |
|
Атомная масса |
|
|
|
|
||
мкмоль |
% |
|
||
|
|
|
||
|
|
|
|
|
Áîð |
2,0 |
2 10–3 |
|
10,82 |
Õëîð |
3,0 |
10–2 |
|
35,46 |
Ñåðà |
30 |
10–1 |
|
32,07 |
Фосфор |
60 |
2 10–1 |
|
30,98 |
Магний |
80 |
2 10–1 |
|
24,32 |
Кальций |
125 |
5 10–1 |
|
40,08 |
Калий |
250 |
1 |
|
39,10 |
|
|
|
|
|
Àçîò |
1000 |
1,5 |
|
14,01 |
|
|
|
|
|
Кислород |
30 000 |
4,5 |
|
16,00 |
|
|
|
|
|
Углерод |
40 000 |
4,5 |
|
12,01 |
|
|
|
|
|
Водород |
60 000 |
6 |
|
1,01 |
|
|
|
|
|
Следует заметить, что качественный состав золы зависит от содержания минеральных элементов в почве и условий внешней среды. Как правило, чем богаче почва и суше климат, тем выше содержание золы в растениях. Любой химический элемент, присутствующий в среде обитания, может быть обнаружен и в растении. Химический состав растения не отображает его потребности в питательных веществах.
Распределение элементов в клеточных структурах весьма различно, что можно проиллюстрировать данными табл. 5.3.
|
Таблица 5.3 |
|
Содержание некоторых элементов в клеточных структурах |
||
|
|
|
Структура |
Элемент |
|
|
|
|
Клеточные стенки |
Кремний, кальций; иногда магний, алюминий |
|
|
|
|
ßäðà |
Кальций, магний, натрий, калий, железо, цинк, медь |
|
|
|
|
Хлоропласты |
Магний, кальций, калий, натрий, марганец, железо, |
|
медь, молибден |
||
|
||
|
|
|
Митохондрии |
Кальций, магний, калий, натрий, железо, медь, цинк |
|
|
|
|
Рибосомы |
Магний, кальций, марганец |
|
|
|
|
Вакуолярный сок |
Калий, натрий, кальций, магний |
|
|
|
|
262
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не являются критерием необходимости.
Хорошо зарекомендовало себя в этом отношении (относительной необходимости) тройное правило Арнона, предложенное еще
â1939 г. Элемент признается необходимым, когда:
1.растение без него не может закончить свой жизненный
öèêë;
2.другой элемент не может заменить функцию изучаемого элемента;
3.элемент непосредственно включен в метаболизм растения. Подвижность разных веществ в почве широко варьирует: вы-
сокоподвижные — калий, натрий, фосфор, сера; среднеподвижные — магний, железо, медь, цинк; относительно подвижные — кальций, магний, бор.
Однако подвижность ионов промежуточной группы зависит от их количества в почве (увеличивается с повышением концентрации).
Принципы подбора природой металлов для построения биомассы до настоящего времени не выяснены, хотя точно известно, что их распространение не было решающим критерием. Так, алюминий и титан очень распространены в природе, однако не являются элементами жизни.
Напротив, молибден, редко встречающийся в природе, очень необходим, например при переносе электронов.
Макро- и микроэлементы
Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10–1—10–2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10–3 % и ниже) — микроэлементами.
Âîäà è ÑÎ2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2Î è ÑÎ2), для галофитов следует добавить Na и Cl.
Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.
Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.
263
В оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений — освещенность соответствующей интенсивности и протяженности.
Минеральные элементы играют определенную роль в обмене веществ растений. Отметим, что в отсутствие наименее важного из этих элементов жизнь была бы невозможна, как и при излишестве любого из них.
Физиологи растений раньше считали, что элементы минерального питания поступают в растения с водой на основе осмоти- ческих и диффузионных ограничений. Однако Д. А. Сабинин показал, что эти процессы независимы:
нет прямой связи между транспортируемым растением количеством воды и количеством солей, поглощаемых из почвенного раствора корневой системой;
наблюдаемое одновременное движение ионов и воды через корневые системы происходит в противоположных направлениях;
механизмы поступления воды и ионов разные. Физиологические функции минеральных элементов и форма
их поступления в клетку приведены в табл. 5.4.
|
|
|
Таблица 5.4 |
|
Физиологические функции минеральных элементов |
||
|
|
|
|
|
Форма, в кото- |
|
Заболевания, свя- |
Элемент |
рой поступает |
Физиологические функции |
занные с недостат- |
|
элемент в клетку |
|
ком элемента |
|
|
|
|
|
Нитрат NO3 , |
Синтез белков, нуклеиновых |
Угнетение роста; |
Àçîò |
кислот и других органических |
сильный хлороз, |
|
аммоний NH4 |
соединений, в частности ко- |
особенно у старых |
|
|
|
ферментов и хлорофилла |
листьев |
|
|
|
|
|
Фосфат HPO4 , |
Синтез нуклеиновых кислот, |
|
|
АТФ и некоторых белков. Вхо- |
Угнетение роста, |
|
Фосфор |
ортофосфат |
дит в состав фосфолипидов |
|
|
H2PO4 |
мембран, играет ключевую |
особенно корней |
|
|
роль в переносе энергии |
|
|
|
|
|
|
|
Синтез белков и многих орга- |
|
|
Сульфат SO24 |
нических соединений, напри- |
Хлороз, например |
Ñåðà |
мер кофермента À, принима- |
пожелтение листь- |
|
|
|
ет участие в ЭТЦ дыхания и |
åâ ÷àÿ |
|
|
фотосинтеза |
|
|
|
|
|
264
Продолжение табл. 5.4
|
Форма, в кото- |
|
Заболевания, свя- |
Элемент |
рой поступает |
Физиологические функции |
занные с недостат- |
|
элемент в клетку |
|
ком элемента |
|
|
|
|
|
|
Осмотический компонент кле- |
Пожелтение и по- |
|
K |
точного сока, активатор или |
бурение листьев с |
Калий |
кофактор многих ферментов, |
краев, преждевре- |
|
|
|
включая АТФазы, основной |
менная гибель рас- |
|
|
потенциалопределяющий ион |
тений |
|
|
|
|
|
|
Стабилизирует структуру ри- |
|
|
Mg2 |
босом. Входит в состав моле- |
Хлороз, |
Магний |
кулы хлорофилла, неспеци- |
||
|
|
фичный кофактор многих фер- |
слабый рост |
|
|
|
|
|
|
ментов |
|
|
|
|
|
|
|
Участвует в формировании |
|
|
|
срединной пластинки (пектат |
|
|
|
кальция) клеточной стенки, |
|
|
|
стабилизатор мембран, кофак- |
|
|
|
тор ряда ферментов, участ- |
|
Êàëü- |
Ca2 |
вующих в гидролизе АТФ и |
Подавление роста, |
фосфолипидов; вероятно, уча- |
снижение прочно- |
||
öèé |
|
ствует в связывании РНК с |
сти хромосом |
|
|
||
|
|
белками в хромосомах, акти- |
|
|
|
ватор хлорных каналов при |
|
|
|
возбуждении, один из мес- |
|
|
|
сенджеров в передаче внутри- |
|
|
|
клеточных сигналов |
|
|
|
|
|
|
|
Входит в состав цитохромов и |
|
|
|
белков с негеминовым желе- |
Сильный хлороз, |
Железо |
Fe2 |
зом, участвующих в фотосин- |
особенно у моло- |
|
|
тезе, фиксации азота и дыха- |
дых листьев |
|
|
нии; катализирует первичные |
|
|
|
реакции синтеза хлорофилла |
|
|
|
|
|
|
|
Принимает участие в фотоли- |
|
|
|
зе воды и восстановлении СО2 |
|
|
|
при фотосинтезе, увеличива- |
|
|
|
ет содержание сахаров и их |
Пятнистость ли- |
Марга- |
Mn2 |
отток из листьев, активирует |
стьев, например |
некоторые дегидрогеназы, де- |
|||
íåö |
|
карбоксилазы и другие фер- |
«серая крапча- |
|
|
тость» у овса |
|
|
|
менты, необходим для функ- |
|
|
|
ционирования нитратредуктаз, |
|
|
|
кофактор РНК-полимеразы, |
|
|
|
ауксиноксидазы |
|
|
|
|
|
265
Продолжение табл. 5.4
|
Форма, в кото- |
|
Заболевания, свя- |
Элемент |
рой поступает |
Физиологические функции |
занные с недостат- |
|
элемент в клетку |
|
ком элемента |
|
|
|
|
|
|
Составная часть алкогольде- |
|
|
|
гидрогеназы (спиртовое бро- |
|
|
|
жение), глутаматдегидрогена- |
Деформация |
Öèíê |
Zn2 |
зы, лактатдегидрогеназы, ще- |
листьев, тормо- |
|
|
лочной фосфотазы и других |
жение роста |
|
|
ферментов, участвует в рас- |
|
|
|
|
|
|
|
щеплении белков, образова- |
|
|
|
нии ростовых веществ |
|
|
|
|
|
|
|
Входит в состав аскорбинок- |
|
|
|
сидазы, тирозина, лактазы, |
|
|
|
моноаминооксидазы, уреазы, |
|
|
|
цитохромоксидазы, галакто- |
|
Ìåäü |
Cu2 |
оксидазы, обеспечивает тер- |
Отмирание побегов |
|
|
минальный перенос электро- |
|
|
|
нов в дыхательной цепи, пе- |
|
|
|
ренос электронов при фото- |
|
|
|
синтезе, влияет на биосинтез |
|
|
|
хлорофилла |
|
|
|
|
|
|
|
Входит в состав нитратредук- |
|
|
|
тазы (восстановление нитра- |
|
|
|
та до нитритов в процессе |
Слабое замедление |
Молиб- |
2 |
синтеза аминокислот), участ- |
|
äåí |
MoO4 |
вует в фиксации атмосферно- |
роста, деформация |
|
побегов |
||
|
|
го азота, в фосфорном обме- |
|
|
|
не, является активатором в |
|
|
|
реакциях аминирования |
|
|
|
|
|
|
|
Обеспечивает нормальное де- |
|
|
|
ление клеток меристемы, уча- |
|
|
|
ствует в процессе транспорта |
Аномальный |
|
|
и обмена углеводов, улучшает |
рост и отмирание |
|
HBO32 |
обеспечение корней О2, âëèÿ- |
верхушек побе- |
Áîð |
ет на белковый, нуклеиновый, |
гов; «сердцевин- |
|
H2BO3 |
фенольный обмены и созрева- |
ная гниль» свек- |
|
|
|
ние семян. Дефицит бора сни- |
лы, растрескива- |
|
|
жает у томатов содержание |
ние стеблей сель- |
|
|
РНК; активирует ростовые |
дерея |
|
|
вещества (рост пыльцевых |
|
|
|
трубок) |
|
|
|
|
|
266
|
|
Окончание табл. 5.4 |
|
|
|
|
|
|
Форма, в кото- |
|
Заболевания, свя- |
Элемент |
рой поступает |
Физиологические функции |
занные с недостат- |
|
элемент в клетку |
|
ком элемента |
|
|
|
|
|
|
Обязателен для некоторых мик- |
|
|
|
роорганизмов, включая сво- |
|
|
|
бодноживущие фиксаторы азо- |
|
|
|
та и симбиотические фикса- |
|
|
|
торы азота бобовых и небобо- |
|
Кобальт |
Co2 |
вых растений. Входит в со- |
Как и при недос- |
|
|
став коэнзимов, содержащих |
татке азота |
|
|
витамин В12, защищает хло- |
|
|
|
рофилл от разрушения в тем- |
|
|
|
ноте, входит в состав окис- |
|
|
|
лительно-восстановительных |
|
|
|
ферментов |
|
|
|
|
|
|
|
Участвует в поддержании ани- |
|
|
|
он-катионного и осмотическо- |
|
Õëîð |
Cl– |
го балансов. Необходим для |
Хлороз |
|
|
фотосинтетических реакций, |
|
|
|
участвующих в выделении ки- |
|
|
|
слорода |
|
|
|
|
|
5.2. Транспорт минеральных веществ в клетку
Минеральные вещества обычно поглощаются из почвы с помощью корней, поступают в небольших количествах и через листья, поэтому внекорневое внесение некоторых микроэлементов стало стандартным сельскохозяйственным методом. Минеральные вещества почти всегда поступают в растения в форме ионов. Ионы сна- чала должны пересечь оболочку и плазмалемму, чтобы попасть в цитоплазму, затем при необходимости пройти через мембрану, окружающую вакуоль (тонопласт) или какую-либо клеточную органеллу, чтобы оказаться в том или ином внутреннем компартменте.
Изучение вопроса транспорта элементов — одна из основных проблем минерального питания. Это предопределяется следующими обстоятельствами: во-первых, суть питания растений состоит в поступлении и включении в метаболизм минеральных элементов в результате обмена между организмом и средой. Во-вторых, изучение процесса транспорта элементов сопряжено с
267
выяснением свойств и функций клеточной оболочки, мембранных образований, связи между клетками и тканями. В-третьих, выяснение вопросов, связанных с транспортом, приближает нас к целенаправленному управлению продуктивностью сельскохозяйственных растений.
Гидратированное состояние ионов
Перенос ионов через мембрану связан с определенными трудностями. Одна из трудностей — наличие гидратированной воды, окружающей ион и значительно увеличивающей его объем.
Рассмотрим одновалентные ионы, в частности катионы Li , Na , K , Rb и Cs : самое легкое ядро у лития, самое тяжелое у цезия. С увеличением массового числа растет и количество электронов, которые окружают ядро, и объем пространства, занимаемого электронными орбиталями. Поскольку плотность электронного облака, окружающего ядро, мала, можно предположить, что ядро более доступно внешним воздействиям. Эта зависимость находит свое отражение в величинах радиусов негидратированных ионов: Li — 0,06; Na — 0,095; K — 0,133; Rb — 0,148; Cs — 0,169 нм.
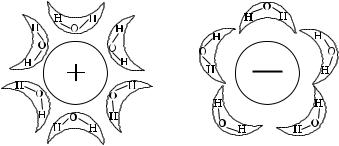
В водных растворах молекулы воды удерживаются около ионов электростатическими силами, источником которых служат заряженные частицы атомного ядра. Чем ближе могут подойти к заряженному атомному ядру молекулы воды, тем сильнее они связываются и тем больше изменяется величина свободной энергии, обусловленная гидратацией. Таким образом, у лития молекулы воды ближе к ядру, и это значит, что в гидратной оболочке лития содержится больше молекул воды. Известно, что молекулы воды — диполи. Около катионов все ближайшие молекулы воды ориентируются отрицательными полюсами внутрь, а около анионов внутрь направлены положительные полюсы молекул воды (рис. 5.1).
Ðèñ. 5.1. Ориентация молекул воды вокруг катиона и аниона
268

Этот внутренний, сильно связанный с ионами структурированный слой молекул воды называют первичной оболочкой.
На некотором отдалении, превышающем толщину первичной оболочки, напряженность электрического поля снижается, что приводит к изменению нормальной ориентации молекул воды. Из-за этого вокруг иона возникает вторичная оболочка (рис. 5.2).
Ðèñ. 5.2. Гидратные оболочки иона в водном растворе
Таким образом, благодаря гидратной оболочке размеры ионов сильно увеличиваются. Между радиусами гидратированных и негидратированных катионов щелочных металлов существует обратная зависимость, т. е. гидратированный ион с меньшим кристаллическим радиусом имеет большие размеры. Радиусы гидратированных ионов трудно вычислить. Подвижность ионов дает представление об их относительных размерах (табл. 5.5).
|
|
|
|
Таблица 5.5 |
|
Подвижность ионов в водных растворах (25 îÑ) |
|||
|
|
|
|
|
Катион |
|
Подвижность, |
Анион |
Подвижность, |
|
ñì2 ñ–1 –1 |
ñì2 ñ–1 –1 |
||
K |
|
7,62 10–4 |
SO24 |
8,27 10–4 |
Na |
|
5,19 10–4 |
Cl– |
7,91 10–4 |
Li |
|
4,01 10–4 |
NO3 |
7,40 10–4 |
В настоящее время общеприняты представления о том, что ионы и различные вещества преодолевают мембрану несколькими способами, основные из которых:
1. Простая диффузия через липидную фазу, если вещество растворимо в липидах (это не касается ионов).
269

2.Облегченная диффузия гидрофильных веществ с помощью липофильных переносчиков (транспортеров).
3.Простая диффузия ионов через гидрофильные поры (например, через ионные каналы).
4.Перенос веществ с участием активных комплексов (насосов).
5.Транспорт веществ путем пиноцитоза в условиях существенных изменений архитектуры мембран.
Что касается движущих сил мембранного транспорта, то разли- чают два механизма: пассивный транспорт и активный транспорт.
Активный транспорт — перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило, в форме АТФ или редокс-цепей.
Пассивный транспорт — перемещение веществ путем диффузии по градиенту электрохимического потенциала (простая и,
âкакой-то мере, облегченная диффузия).
Пассивный транспорт. Простая диффузия
Чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на некоторых физикохимических закономерностях, определяющих движение ионов в растворе и мембране.
Согласно первому закону Фика, поток вещества (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dõ в точке õ в данный момент времени
Ô D |
dC |
. |
(5.1) |
|
|||
|
dx |
|
|
Знак минуса в правой части уравнения означает, что, если градиент положителен, т. е. концентрация при увеличении õ возрастает, диффузионный поток направлен в противоположную сторону.
Однако при описании диффузии на большие расстояния в непрерывной системе необходимо определить зависимость концентрации не только от расстояния (x), но и от времени (t). Поэтому целесообразно преобразовать первый закон Фика в дифференциальные уравнения в частных производных, которые обычно называют вторым законом Фика. В одномерном случае этот закон выражается следующим образом:
dC |
D |
d2C |
. |
(5.2) |
|
|
|||
dt |
|
dx2 |
|
|
270

Таким образом, скорость изменения концентрации пропорциональна второй производной от концентрации на координате õ. Из этого закона получаем важное соотношение
x 
 4Dt. (5.3)
4Dt. (5.3)
Скорость прохождения иона через мембрану при простой диффузии в сильной степени зависит от его поведения в самой мембране (липидном бислое). Важные параметры — активность и подвижность данного иона в самой мембране. Так как действительный градиент концентрации в мембране неизвестен, то силой, определяющей движение молекул, считают средний градиент
DC |
|
C |
|
C â Ñ í |
(5.4) |
|
|
|
|
|
|
. |
|
|
|
|
||||
dx |
|
x |
|
õ |
|
|
Если обозначить через S количество вещества, которое входит в клетку, то dS/dt характеризует скорость поступления этого вещества. Поток Ф — это скорость поступления вещества в пересче- те на единицу площади (A), т. е. (1/А) (dS/dt).
На поверхности мембраны в растворе концентрация веществ может отличаться. Учитывая коэффициент распределения K и все другие соображения, первый закон Фика можно записать следующим образом:
1 |
|
dS |
|
C í Ñ â |
(5.5) |
|
|
|
|
DK |
|
, |
|
A |
|
|
|
|||
|
dt |
|
õ |
|
||
ãäå dS/dt — положительная величина, если вещество поступает в клетку, т. е. если концентрация вещества в среде больше, чем его концентрация в клетке.
В (5.5) нам неизвестны ни коэффициент распределения, ни коэффициент диффузии, ни действительная величина õ, поэтому эти три параметра целесообразно заменить одним — коэффициен-
том проницаемости |
|
|
|
P |
DK |
(ñì/ñ). |
(5.6) |
|
|||
|
x |
|
|
Коэффициент проницаемости мембраны — это сумма молей вещества, которая прошла через 1 см2 мембраны в единицу времени, при условии разности концентраций по обеим сторонам мембраны, составляющей 1 моль/см3.
271