

Вищі полісахариди
Вищі полісахариди – високомолекулярні речовини, молекули яких містять від декількох десятків до багатьох тисяч залишків моносахаридів. Вивчення цих полісахаридів ускладнюється труднощами їх очищення і одержання у чистому вигляді, оскільки в клітині вони звичайно зв'язані з іншими вуглеводами, білками, мінеральними речовинами. При вивченні вищих полісахаридів використовують метод введення в їхні молекули різних радикалів (–СН3, –СОСН3), що дозволяє легше очистити отриманий препарат, вивчити його будову і зв'язки між залишками моносахаридів. Використовують також метод м'якого кислотного або ферментного гідролізу полісахаридів, що дозволяє встановити склад їх мономерів.
Вищі полісахариди поділяють на дві групи: гомо- і гетерополісахариди. Молекули гомополісахаридів побудовані із залишків одного цукру. Так, із залишків глюкози складаються крохмаль, целюлоза, глікоген, калоза, ліхенін, а із залишків фруктози – інулін, леван. До складу молекул гетерополісахаридів входять залишки різних моносахаридів та їхніх похідних – геміцелюлози, гумі, слизи.
Молекули вищих полісахаридів можуть бути як лінійними, так і розгалуженими. Усі полісахариди цієї групи не мають відновних властивостей, тому що у зв'язках між залишками цукрів беруть участь глікозидні гідроксили.
Крохмаль – головний запасний полісахарид рослин, який відкладається у клітинах органів у вигляді крохмальних зерен (насіння, плоди, корені, бульби, кореневища, стебла). Особливо багато крохмалю в насінні рису (60-80 %), кукурудзи (65-75 %), пшениці (60-70 %), менше у бульбах картоплі (12-22%).
Крохмальні зерна не розчиняються у воді, а тільки набрякають. Якщо завись крохмальних зерен у воді поступово нагрівати, і о може бути досягнута температура, при якій крохмаль утворює дуже в'язкий колоїдний розчин – крохмальний клейстер. Температура клейстеризації неоднакова для крохмалю різних рослин. Крохмаль картоплі клейстеризується при температурі 55-65 °С, кукурудзи – при 64-71 °С, пшениці – при 60-80 °С, рису – при 70-80 °С.
Характерною властивістю крохмалю є його здатність давати синій колір при додаванні розчину йоду в йодистому калії.
Крохмаль на 96-98 % складається з вуглеводів. Інше становлять домішки – мінеральні речовини і жирні кислоти.
Вуглеводна частина крохмалю складається з двох полісахаридів: амілози і амілопектину.
Амілоза являє собою довгий нерозгалужений ланцюг із залишків α-глюкози, поєднаних (α1→4)-зв'язками. Ланцюги амілози утворюють спіраль, кожен виток якої має 6 залишків глюкози. У молекулі амілози звичайно містяться декілька паралельно розташованих спіралей.
Молекулярна маса амілози від 100 000 до 1 000 000. При дії на амілозу розчину йоду його молекули вбудовуються у витки спіралі, утворюючи комплексну з полісахаридом хімічну сполуку, що має синє забарвлення.
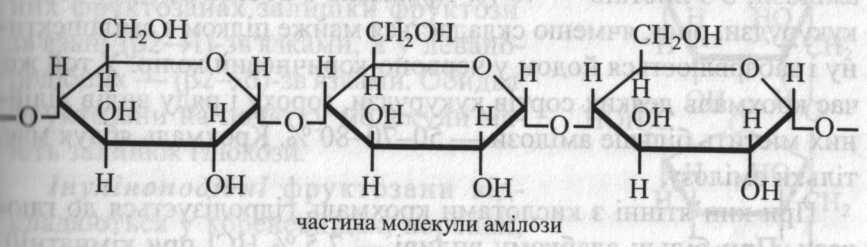
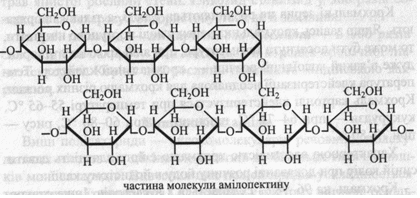
Амілопектин побудований із залишків α-глюкози, поєднаних (α1→4)- і (α1→6)-зв'язками, тобто молекула амілопектину має розгалужену структуру. Точки розгалуження зустрічаються через 25-30 глюкозних залишків.
Зовнішні ланцюги амілопектину, якщо вони мають більше шести залишків глюкози, можуть утворювати витки спіралі. Молекулярна маса амілопектину вища, ніж у амілози – від одного до декількох мільйонів.
Амілопектин з йодом дає червоно-фіолетове забарвлення, що є результатом адсорбції молекул йоду на полісахариді без утворення хімічних зв'язків.
Як правило, крохмаль містить 10-30 % амілози і 70-90 % амілопектину. Однак це співвідношення може змінюватися в залежності від виду або сорту рослин, а також від органа, з якого отриманий крохмаль. Так, у крохмалі з бульб картоплі міститься 22 % амілози, а з листків – 46 %. Крохмаль з воскоподібних сортів кукурудзи, рису, ячменю складається майже цілком з амілопектину і забарвлюється йодом у червоно-коричневий колір. У той же час крохмаль деяких сортів кукурудзи, гороху і ряду видів лілійних містить більше амілози – 50-70-80 %. Крохмаль яблук має тільки амілозу.
При кип'ятінні з кислотами крохмаль гідролізується до глюкози. При більш слабкому впливі – 7,5 % НС1 при кімнатній температурі протягом 7 днів – утворюється так званий «розчинний крохмаль», який часто застосовується у лабораторіях.
Крохмаль має широке практичне використання:
-
він складає велику частину їжі людини (хліб, крупи, овочі, фрукти, кондитерські вироби) і кормів сільськогосподарських тварин;
-
використовується у багатьох галузях харчової промисловості;
-
з нього одержують глюкозу, спирт, клей, пластмаси;
-
його використовують у текстильній промисловості.
Глікоген (тваринний крохмаль) міститься в тканинах тварин, у грибах, у ряді рослин (зерно цукрової кукурудзи). У дріжджах глікогену до 40 %. За складом і структурою молекул він близький до амілопектину, але відрізняється від нього більшою розгалуженістю і компактністю молекули, тому що точки розгалуження у глікогену розташовані через 10-18 залишків глюкози. Молекулярна маса його також вища, ніж в амілопектину. Глікоген розчиняється у гарячій воді, створюючи кооїдний розчин, що опалесціює.
Глікоген відіграє важливу роль у вуглеводному обміні тварин, грибів, дріжджів, при спиртовому бродінні.
Ф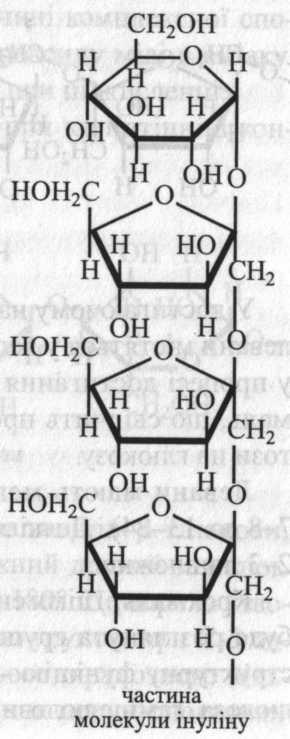 руктозани
– полімерні сполуки, що складаються із
залишків β-фруктози. Існує два типи
фруктозанів – інуліноподібні
й
леваноподібні.
Вони
розчиняються у воді й містяться в
клітинному соку вакуолі. В інуліноподібних
фруктозанах залишки фруктози зв'язані
(β2→1)-зв'язками,
а у леваноподібних – (β2→6)-зв'язками.
Обидва фруктозани на початку молекули
мають залишок глюкози.
руктозани
– полімерні сполуки, що складаються із
залишків β-фруктози. Існує два типи
фруктозанів – інуліноподібні
й
леваноподібні.
Вони
розчиняються у воді й містяться в
клітинному соку вакуолі. В інуліноподібних
фруктозанах залишки фруктози зв'язані
(β2→1)-зв'язками,
а у леваноподібних – (β2→6)-зв'язками.
Обидва фруктозани на початку молекули
мають залишок глюкози.
Інуліноподібні фруктозани відкладаються у кореневищах, коренях, бульбах рослин із родини айстрових, дзвоникових (топінамбур, жоржини, цикорій, артишоки, кульбаба). Головним фруктозаном цієї групи є інулін, він містить 30-35 залишків фруктози.
Інулін часто супроводжується більш низькомолекулярними фруктозанами. Вони утворюють гомологічний ряд, на початку якого знаходиться дисахарид із двох залишків фруктози – β-левулін, а наприкінці – інулін. З інуліну шляхом кислотного гідролізу одержують фруктозу.
Леваноподібні фруктозани характерні для однодольних, у тому числі для рослин з родин лілійних і злакових. Левани містяться в листках, стеблах, коренях і насінні. У злаків левани функціонують як тимчасові запасні полісахариди.
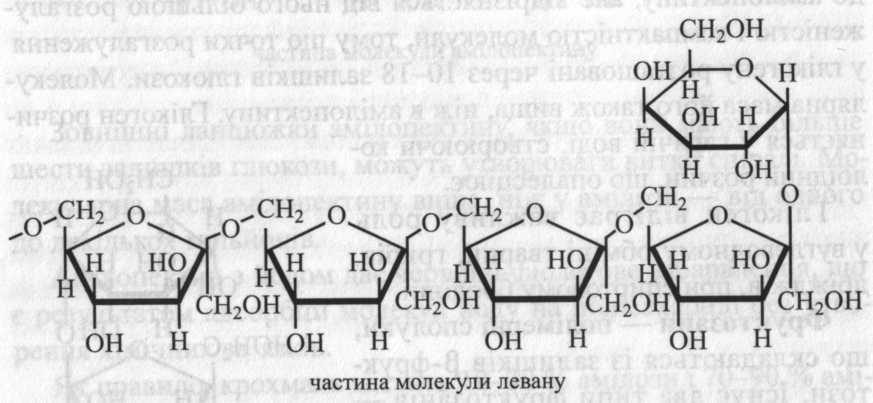
У достигаючому насінні злаків (жито, пшениця, овес, ячмінь) левани містяться у великій кількості (до 30 % на суху речовину), у процесі достигання вони поступово перетворюються на крохмаль, що свідчить про легкість перетворення в рослинах фруктози на глюкозу.
Левани мають менше залишків фруктози, ніж інулін (від 7-8 до 13-24). Деякі левани утворюють слабке розгалуження із 2-3 ланцюжків.
Крохмаль, глікоген, інулін – це запасні полісахариди. Далі буде розглянута група полісахаридів, що виконують у рослині структурну функцію – компонентів клітинної оболонки. Це целюлоза, геміцелюлози і пектини.
Целюлоза (клітковина) – вищий полісахарид, основний компонент клітинної оболонки, що утворює її каркас.
У рослинах клітковина завжди пов'язана з іншими речовинами – геміцелюлозами, лігніном, пектином, ліпідами, смолами тощо. Для видалення цих сполук з клітковини вихідну речовину обробляють сумішшю NaHSO3 і Н2SO3. При цьому супутні речовини переходять у розчин, і залишається чиста целюлоза – біла волокниста речовина.
Основним джерелом одержання целюлози є волокна бавовнику, луб'яні волокна прядив'яних рослин (льону, коноплі, джута, рамі), а також солома і деревина. Бавовняне волокно містить 95-98 % клітковини, лляне – 80-90 %, деревина – 40-50 %.
Целюлоза не розчиняється у воді, а тільки набрякає. Вона є дуже стійкою речовиною: не змінюється під дією слабких кислот і лугів навіть при кип'ятінні, не розчиняється в більшості звичайних розчинників. Клітковина розчиняється тільки в реактиві Швейцера [Сu(NH3)4]•(ОН)2 – розчині комплексної сполуки, що утворюється при розчиненні гідроксиду міді в аміаку. З цього розчину клітковина випадає в осад при підкисленні.
Гідролізується клітковина до глюкози при кип'ятінні з концентрованою НСl або Н2SO4.
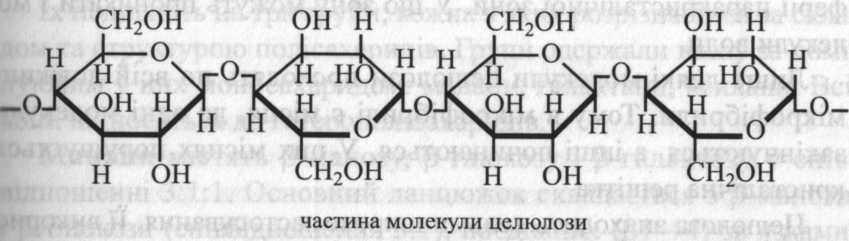
Молекула
целюлози побудована із залишків
β-глюкози, з'єднаних (β1→4)-зв'язками.
Причому кожний другий залишок і глюкози
повернений щодо попереднього на 180°. Це
сприяє утворенню водневого зв'язку між
гідроксильною групою при С-3 одного
глюкозного залишку і киснем сусіднього
піранозного кільця. Це перешкоджає
обертанню розташованих поруч глюкозних
залишків навколо їхніх глікозидних
зв'язків і унеможливлює утворення
спіралі, подібної до амілози. У результаті
утворюється жорстка лінійна структура,
у якій усі піранозні кільця розташовуються
в одній площині. Це дуже важливо для
речовини, що утворює каркас клітинної
оболонки та забезпечує її міцність.
Целюлоза являє собою суміш гомологічних
молекул з м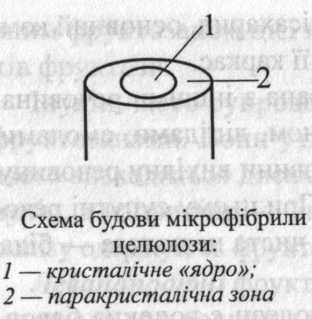 олекулярною
масою від декількох сотень тисяч до
декількох мільйонів.
олекулярною
масою від декількох сотень тисяч до
декількох мільйонів.
У клітинних стінках молекули целюлози зібрані у пучки – мікрофібрили, у яких вони розташовані паралельно одна до одної та пов'язані водневими зв'язками. У поперечному розрізі мікрофібрила має овальну форму.
У центрі мікрофібрили молекули (їх приблизно 50) складають так зване «ядро». У ньому молекули розташовані упорядковано, утворюючи кристалічну решітку. Навколо «ядра» приблизно 100 молекул також розташовані паралельно, але менш упорядковано. Це паракристалічна ділянка. У ній міститься деяка кількість молекул матриксу оболонки, частіше геміцелюлоз. Причому вміст їх збільшується у напрямку від «ядра» до периферії паракристалічної зони. У цю зону можуть проникати і молекули води.
Лише деякі молекули целюлози проходять по всій довжині мікрофібрили. Тому в мікрофібрилі є місця, де одні молекули закінчуються, а інші починаються. У цих місцях порушується кристалічна решітка.
Целюлоза знаходить вельми широке застосування. її використовують у целюлозно-паперовій промисловості. У текстильній промисловості з неї виготовляють бавовняні і лляні тканини. Целюлозу розчиняють у мідно-аміачному розчині, а потім його продавлюють крізь тонке сито у кисле середовище, де вона випадає в осад у вигляді дуже тонких ниток. Із них виробляють натуральний шовк – віскозу. Однак у ньому нитки складаються не з білка, а з вуглеводу – целюлози.
У молекули целюлози вводять різні радикали та одержують метил-, ацетил- або нітроцелюлозу, які є сировиною для багатьох галузей хімічної промисловості. З них виготовляють штучне волокно і тканини, шкіру, пластмаси, фарби, лаки, вибухові речовини тощо.
Целюлозу почали використовувати для виготовлення целюлозобетону, який одержують змішуванням цементу з волокнистою рослинною сировиною – рисовою соломою, бамбуком, сезалем, кокосовими волокнами, коноплями, джутом. Із суміші готують панелі, що мають усі якості залізобетону, але дешевші, легші й зручніші у житловому будівництві.
Геміцелюлози (напівклітковина) – це група вищих полісахаридів, що поряд з целюлозою входять до складу клітинних оболонок, створюючи їх матрикс. У насінні вони можуть частково використовуватися в якості запасних речовин. Досить високий вміст цих сполук у насінні, соломі, деревині, кукурудзяних качанах, висівках.
Геміцелюлози нерозчинні у воді, але розчиняються в лугах і гідролізуються кислотами легше, ніж клітковина. При кислотному гідролізі геміцелюлоз утворюються глюкоза, галактоза, маноза, ксилоза, L-арабіноза й уронові кислоти. Молекулярна маса геміцелюлоз складає декілька десятків тисяч.
Їх поділяють на три групи, кожна з яких розрізняється за складом та структурою полісахаридів. Групи одержали назву за домінуючим у них моносахаридом: манани, галактани, ксилани. Всі вони відносяться до гетерополісахаридів.
Манани містять β-манозу, β-глюкозу і β-галактозу у співвідношенні 3:1:1. Основний ланцюг складається з β-манози і β-глюкози (співвідношення 3:1), поєднаних (β1→4)-зв'язками. До деяких залишків манози приєднані (β1→6)-зв'язками поодинокі залишки галактози. Ряд залишків манози ацетильовані за С-2, рідше за С-3. Такі манани зустрічаються, наприклад, у деревині хвойних і папоротей.
Галактани в основному ланцюгу мають залишки β-галактози, поєднані (β1→3)-зв'язками. До залишків галактози (β1→6)-зв'язками приєднуються залишки дисахаридів з двох β-галактоз або двох L-арабіноз. Галактани звичайно містяться в деревині.
Ксилани різноманітніші за структурою, у них переважає ксилоза, незалежно від того чи складає вона головний ланцюг, чи розгалуження. Існує декілька груп ксиланів.
-
Основний ланцюг складається з β-ксилози, (β1→4)-зв'язки, 7 з кожних 10 залишків ксилози ацетильований за С-3, рідше за С-2. До деяких ксилозних залишків приєднана галактуронова кислота (α1→2)-зв'язком, яка містить метоксильну групу біля С-4. Іноді такі ксилани у відгалуженнях мають α-L-арабінозу за (α1→3)-зв'язком. Подібні ксилани зустрічаються у хвойних і папоротей.
-
Інший вид ксиланів містить ксилозу головним чином у бічних гілках. Такі ксилани характерні для клітинних стінок дводольних. Вони поділяються на такі підгрупи:
а) основний ланцюг складається із залишків глюкози, (β1→4)-зв'язок. Відгалуження – поодинокі залишки ксилози, (α1→6)-зв'язок;
б) деякі залишки глюкози основного ланцюга мають розгалуження у вигляді трисахариду з α-ксилози, α-L-фукози, α-галактози. Зв'язок з основним ланцюжком через ксилозу, (α1→6);
в) деякі залишки глюкози основного ланцюга мають два відгалуження: зазначений вище трисахарид і L-арабіноза, (α1→2)-зв'язок. Іноді цей ксилан пов'язаний через глюкозу з пектином.
Пектини – компоненти матриксу первинних клітинних оболонок. Звичайно вони також є міжклітинною речовиною, що утворює серединну пластинку, яка склеює стінки сусідніх клітин. Вміст пектинів у клітинній оболонці невеликий – менше 5 %.
Характерною властивістю пектинів є здатність утворювати гелі при низькій концентрації. Тому їх використовують у харчовій промисловості як желеутворювальні агенти при виготовленні фруктових желе, мармеладу, пастили, джемів, фруктових карамельних начинок і в домашніх умовах при готуванні варення і повидла.