

Принципы эндокринной регуляции
Пептидные гормоны связываются с рецепторами, встроенными в клеточную мембрану. Взаимодействие гормона с рецептором активирует внутриклеточный сигнальный путь. При действии на клетку различных внеклеточных сигналов эти пути различаются. Если клетка является мишенью нескольких гормонов, то она может различать действие конкретного гормона, который активирует определённый внутриклеточный сигнальный путь. Однако некоторые гормоны действуют на одну и ту же клетку-мишень, активируя один и тот же сигнальный путь.
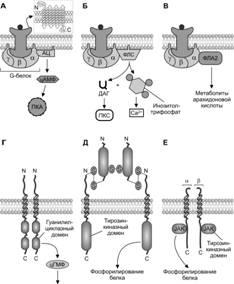
Рецепторы и внутриклеточные эффекторы пептидных гомонов. Молекула рецепторов паратиреокрина, АДГ, тиролиберина имеет сходное строение и содержит семь трансмембранных участков; внутриклеточный домен молекулы рецептора связан с комплексом субъединиц G–белка. В случае паратиреокрина -субъединица данного комплекса взаимодействует с аденилатциклазой (AС), активация которой приводит к увеличению уровня цАМФ в цитозоле с последующим фосфорилированием протеинкиназы А (PKA). -субъединица G–белка, связанного с рецептором АДГ, взаимодействует с фосфолипазой С, которая активирует образование инозитолтрифосфата (IP3) и диацилглицерола (DAG). При активации рецепторов инозитолтрифосфата из цистерн гладкой эндоплазматической сети освобождается Ca2+. Рецептор тиролиберина через -субъединицу активирует фосфолипазу А2 (PLA2) и метаболизм арахидоновой кислоты в клетке-мишени. Молекулы рецепторов предсердного натрийуретического фактора (атриопептина), инсулина и гормона роста имеют сходную структуру, состоят из двух параллельно расположенных субъединиц и соответственно двух трансмембранных участков. Во внутриклеточный С-конец каждой субъединицы встроен в рецепторе натрийуретического фактора гуанилатциклазный домен, а в рецепторе инсулина — тирозинкиназный домен. Внутриклеточная часть субъединиц молекулы рецептора гормона роста связана с киназой Janus (JAK) или другими киназами. Активация рецептора натрийуретического фактора приводит к увеличению в клетке-мишени содержания второго посредника цГМФ. Эффекты инсулина и гормона роста опосредуются через фосфорилирование упомянутыми киназами многих эффекторных белков.

Рецепторы катехоламинов. 1- и 2-адренорецепторы и рецепторы дофамина DA-1 связаны с Gs, который активирует аденилатциклазу (AC), что приводит к увеличению содержания цАМФ в цитозоле. 2-Адренорецепторы, рецепторы дофамина DA-2 взаимодействуют с GI, который подавляет активность аденилатциклазы. 1-Адренорецептор взаимодействует с белком Gq, который активирует фосфолипазу С (PLC) с последующим конвертированием фосфоинозитидов в инозитолтрифосфат (IP3) и диацилглицерол (DAG).
Варианты регуляции активности клеток-мишеней
В зависимости от расстояния от продуцента гормона до клетки–мишени различают эндокринный, паракринный и аутокринный варианты регуляции.

Варианты воздействия лигандов на клетки–мишени.
Эндокринная, или дистантная регуляция. Секреция гормона происходит во внутреннюю среду, клетки–мишени могут отстоять от эндокринной клетки сколь угодно далеко. Наиболее яркий пример: секреторные клетки эндокринных желёз, гормоны из которых поступают в систему общего кровотока.
Паракринная регуляция. Продуцент биологически активного вещества и клетка–мишень расположены рядом, молекулы гормона достигают мишени путём диффузии в межклеточном веществе. Например, в париетальных клетках желёз желудка секрецию H+ стимулируют гастрин и гистамин, а подавляют соматостатин и простагландины, секретируемые рядом расположенными клетками.
Аутокринная регуляция. При аутокринной регуляции сама клетка–продуцент гормона имеет рецепторы к этому же гормону (другими словами, клетка–продуцент гормона в то же время является собственной мишенью). В качестве примера приведём эндотелины, вырабатываемые клетками эндотелия и воздействующие на эти же эндотелиальные клетки; T–лимфоциты, секретирующие интерлейкины, имеющие мишенями разные клетки, в том числе и T–лимфоциты.
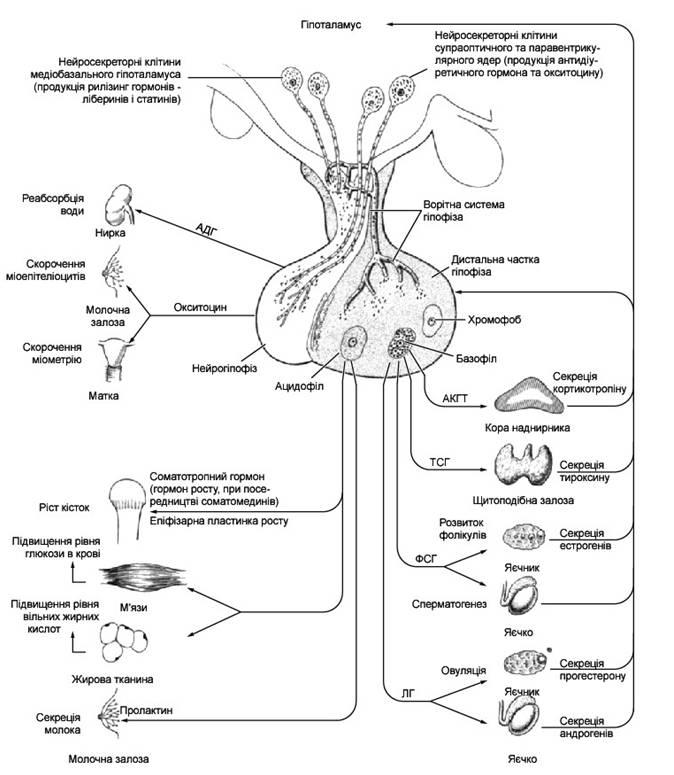
Механизм действия гормонов
Молекула гормона, которая циркулирует с током крови или лимфы, «узнает» свой рецептор на поверхности той или другой клетки-цели. Определяющую роль в этом высокоспецифическом узнавании имеет соответствие стереохимии активного центра молекулы гормона и конфигурации его рецептора. Здесь, как и в ряде других жизненно важных процессов организма, действует принцип соответствия «ключа» (гормон) к «замку» (рецептор плазмолемы). Связывание гормона с рецептором вызывает конформационные (объемно-пространственные) изменения молекулы рецептора, что, в свою очередь, влияет на ферментные системы клетки, в частности на аденилат-циклазную систему. При этом фермент аденилатциклаза предопределяет превращение АТФ в циклический аденозинмонофосфат (цАМФ). Молекулы последнего играют роль универсального стимулятора внутрицитоплазматических ферментов клетки. Характерным является то, что эффект действия гормонов может проявляться не только в усилении, но и в угнетении деятельности клеток и их систем. Заметим, что за открытие универсальной роли цАМФ в механизме действия гормонов американский исследователь Е. Сазерленд в 1971 г. удостоен Нобелевской премии.
В основе взаимодействия между отдельными звеньями эндокринной системы, а также между эндокриноцитами и клетками-целями лежит принцип обратной связи. Влияние того или другого гормона на клетку-цель приводит к усилению продуцирования ею определенных химических веществ. Повышение концентрации последних во внутренней среде организма становится своеобразным сигналом к угнетению деятельности эндокриноцита. Напротив, уменьшение концентрации гормона в крови или лимфе является стимулом синтетической деятельности эндокриноцита. Принцип обратной связи сохраняет свою силу и в случае подавляющего (ингибиторного) влияния гормона на орган-цель.
Все эндокринные железы имеют ряд общих черт строения :
1. В их составе отсутствуют выводные протоки .
2 .се они имеют хорошо развитую сосудистую сеть, особенно микроциркуляторного русла (капилляры висцерального типа с фенестрованным эндотелием и сплошной базальной мембраной) .
3. Клетки эндокринных органов образуют характерные скопления в виде фолликулов (пузырьков)б или трабекул (перекладинб или частиц. Тоесть, паренхима этих желез имеет соответствующие структурные или морфофункциональные единицы - фолликулы, трабекулы , дольки.
4. В эндокриноцитах ( клетках - производителях гормонов) , как правило , можно обнаружить специфические гранулы, в которых накапливается биологически активное вещество .
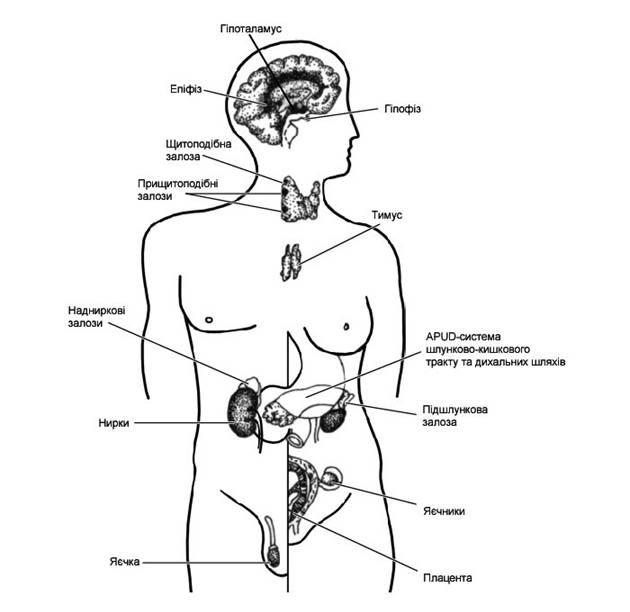
Условно среди элементов эндокринной системы организма различают четыре группы составляющих: центральные , периферийные , органы со смешанными функциями и диссоциированные эндокринные клетки.Условно среди элементов эндокринной системы организма различают четыре группы составляющих частей. К первой группе — центральным органам эндокринной системы — принадлежат гипоталамус, гипофиз и эпифиз. Эти органы тесно связаны с органами центральной нервной системы и координируют деятельность всех других звеньев эндокринной системы. Вторая группа — периферические эндокринные органы — включает щитовидную, прищитовидные и надпочечные железы. Это сугубо эндокринные железы, которые осуществляют многоплановое влияние на организм, усиливая или ослабляя обменные процессы. Третья группа элементов включает органы, которые совмещают выполнение эндокринной функции с рядом других. Это поджелудочная железа, половые железы (яичко, яичник), почки, плацента и тому подобное. В организме человека есть также большая группа клеток — так называемая дисоциированная эндокринная система — которые образуют четвертую группу элементов эндокринной системы.