

БХ - 3 семестр / ()Лекции / ()Общие / Углеводы / Полисахариды
.docПолисахариды, потребляемые организмом в результате метаболизма, поступают частично с пищей (амилоза + амилопектин= крахмал), частично имеют эндогенное происхождение (гликоген). Суточная потребность организма в углеводах- 400-500 г. Источник- мучные изделия, крупы, злаки, фрукты и мед. На начальных этапах происходит расщепление полисахаридов до олиго- и моносахаридов под воздействием ряда ферментов (амилаза, мальтаза, изомальтаза и некоторые другие). Переваривание начинается в тонком кишечнике под воздействием фермента АМИЛАЗЫ поджелудочной железы:
крахмал (АМИЛОЗА) --> МАЛЬТОЗА -->ГЛЮКОЗА
В конечном итоге образуется глюкоза из всех вышеперечисленных полисахаридов. Образовавшаяся глюкоза претерпевает превращения по нескольким направлениям.
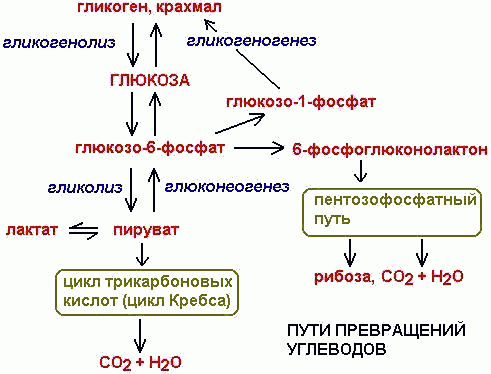
1) ФОСФОРИЛИРОВАНИЕ ГЛЮКОЗЫ до Г-6-Ф:

Г-6-Ф непроходим через мембраны. Реакция катализируется ГЕКСОКИНАЗОЙ во всех клетках и ГЛЮКОКИНАЗОЙ в печени. Глюкокиназа не ингибируется Г-6-Ф и в печени накапливается значительное количество Г-6-Ф, предупреждая значительное повышение глюкозы в крови на пике пищеварения.
ПРЕВРАЩЕНИЯ Г-6-Ф
1) Под действием ФОСФАТАЗЫ обратное превращение в глюкозу и фосфат.
2) ФОСФОГЛЮКОМУТАЗА катализирует превращение Г-6-Ф в Г-1-Ф (промежуточный продукт синтеза ГЛИКОГЕНА):
 При
взаимодействии с УТФ глюкозо-1-фосфат
образует УДФ-глюкозу, которая далее
переносит остаток глюкозы на растущую
цепь гликогена. Регенерация УТФ происходит
в результате реакции УДФ с молекулой
АТФ.
При
взаимодействии с УТФ глюкозо-1-фосфат
образует УДФ-глюкозу, которая далее
переносит остаток глюкозы на растущую
цепь гликогена. Регенерация УТФ происходит
в результате реакции УДФ с молекулой
АТФ.

3) Г-6-Ф-ДЕГИДРОГЕНАЗА превращает Г-6-Ф в ГЛЮКОНОЛАКТОН:

который далее превращается по ПЕНТОЗОФОСФАТНОМУ ПУТИ.
4) Г-6-Ф изомеризуется во ФРУКТОЗО-6-ФОСФАТ и вовлекается в превращения вплоть до углекислого газа и воды (основной путь катаболизма).
ГЛИКОЛИЗ- АНАЭРОБНЫЙ РАСПАД ГЛЮКОЗЫ
Является ПЕРВЫМ ЭТАПОМ распада глюкозы. В процессе гликолиза молекула глюкозы распадается до двух молекул пировиноградной кислоты- пирувата. При этом образуется две молекулы восстановительного эквивалента- NADH, и две молекулы АТФ.
(Обратное превращение пирувата в глюкозу называется глюконеогенезом. Часть стадий глюконеогенеза совпадает со стадиями гликолиза. Это обратимые стадии для обоих процессов. Необратимые стадии гликолиза заменены в глюконеогенезе на превращения, катализируемые другими ферментами. Стадии гликолиза приведены в правом столбце таблицы, глюконеогена- в левом. В центральном столбце показаны интермедиаты превращений и обозначен номер стадии).
|
ГЛЮКОНЕОГЕНЕЗ |
интермедиаты |
ГЛИКОЛИЗ |
|
||||||
|
Катализаторы стадий ГЛЮКОНЕОГЕНЕЗа (обходные пути) |
ГЛЮКОЗА |
Катализаторы стадий гликолиза (в том числе общие с глюконеогенезом) |
|
||||||
|
Г-6-фосфатаза (с высвобождением Фн) |
1- необратимая (ингибитор- Г-6-Ф) |
Глюкокиназа (печень) Гексокиназа (др.клетки) (с затратой АТФ) (с высвобождением АДФ, Фн) |
|
||||||
|
ГЛЮКОЗО-6-ФОСФАТ |
|
||||||||
|
|
2 |
Г-6-Ф-изомераза |
|
||||||
|
ФРУКТОЗО-6-ФОСФАТ |
|
||||||||
|
Фруктозо-1,6-бифосфатаза (с высвобождением Фн) |
3-необратимая (активаторы- АМФ, Ф-2,6-ДФ, ингибиторы- АТФ, цитрат, Н+) |
Фосфофрукто-1-киназа (с затратой АТФ) |
|
||||||
|
ФРУКТОЗО-1,6-ДИФОСФАТ |
|
||||||||
|
|
4 |
Альдолаза |
|
||||||
|
2 х ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТ |
|
||||||||
|
(с затратой 2 NADH) |
5 |
(с затратой 2NAD+, Фн) Г-3-Ф-дегидрогеназа (с высвобождением 2NADH) |
|
||||||
|
2 х 1,3-ДИФОСФОГЛИЦЕРАТ |
|
||||||||
|
|
6 |
|
|
||||||
|
2 х 2,3-ДИФОСФОГЛИЦЕРАТ |
|
||||||||
|
(с затратой 2АТФ) |
7 |
Фосфоглицераткиназа (с высвобождением 2АТФ) |
|
||||||
|
2 х 3-ФОСФОГЛИЦЕРАТ |
|
||||||||
|
|
8 |
Фосфоглицератмутаза |
|
||||||
|
2 х 2-ФОСФОГЛИЦЕРАТ |
|
||||||||
|
|
9 |
Енолаза +Н2О |
|
||||||
|
|
2 х ФОСФОЕНОЛПИРУВАТ (ФЕП) |
|
|
||||||
|
ФЕП-карбоксикиназа (с затратой 2АТФ), (с высвобождением 2СО2) |
10б |
|
|||||||
|
ОКСАЛОАЦЕТАТ |
10-Необратимая |
Пируваткиназа (с высвобождением 2АТФ) |
|
||||||
|
Пируваткарбоксилаза (с затратой 2АТФ, 2СО2) |
10а |
|
|
|
|||||
|
2х ПИРУВАТ (ПВК) |
|
||||||||
|
|
11 (ответвление цепи) |
Лактатдегидрогеназа (с затратой 2NADН) (с высвобождением 2NAD+) |
|
||||||
|
затрачено 6 АТФ, 2 NADH |
2 х ЛАКТАТ |
|
|
||||||
|
|
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС |
получено 2 АТФ, 2 NADH |
|
||||||
|
|
|
|
|
|
|||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
|
|
||||||||
|
№ стадии |
Катализатор гликолитического пути |
Активатор |
Ингибитор стадии |
|
1 |
глюкокиназа (печень), гексокиназа (мышцы) |
|
Г-6-Ф (для гексокиназы) |
|
2 |
фосфоглюкоизомераза |
|
|
|
3 |
6-фосфофрукто-1-киназа |
АМФ, фруктозо-2,6-дифосфат |
АТФ, цитрат, Н+ |
|
4 |
Ф-1,6-ДФ-альдолаза |
|
|
|
5 |
глицеральдегид-3-фосфат-дегидрогеназа |
|
|
|
6 |
фосфоглицерат-мутаза |
|
|
|
7 |
фосфоглицерат-киназа |
|
|
|
8 |
фосфоглицерат-мутаза |
|
|
|
9 |
енолаза |
|
|
|
10 |
пируват-киназа |
фруктозо-1,6-дифосфат |
АТФ, аланин |
|
11 |
лактатдегидрогеназа |
|
|

Катализаторами стадий, эффекторами стадий превращений в гликолизе являются:
В анаэробных условиях молочная кислота конечный продукт, накапливается в мышцах, оттуда поступает в кровь. В печени и в самих мышцах 1/5 часть ее окисляется до СО2 и Н2О.
Гликолиз протекает в цитоплазме. Иницииаторы гликолиза- совершаемая клетками работа, соответственно, накапливание АМФ. Интенсивная мышечная деятельность усиливает гликолиз, накапливается значительное количество молочной кислоты, в результате чего возникает боль в мышцах. Гликолиз должен сопровождаться удалением восстановительных эквивалентов (NADH), которые одним из челночных механизмов переносят электроны внутрь митохондрий, где те используются в дыхательной цепи (сопряженном фосфорилировании).
Энергетическая эффективность ГЛИКОЛИЗА- 35%, в результате гликолиза образуется (нетто): 2 молекулы АТФ и 2 молекулы NADH, из которых в дыхательной цепи синтезируется 6 молекул АТФ (итого- 8 молекул АТФ). Биологическое значение- образование энергии в моменты максимальной потребности в ней и при кислородном голодании.
Регуляция гликолиза
Влияние ГЛЮКАГОНА. Глюкагон взаимодействует с рецепторами на клеточной мембране. Рецепторы глюкагона связаны с работой аденилатциклазы, которая в результате присоединения глюкагона к рецептору синтезирует медиаторы- 3,5-ц-АМФ. ц-АМФ активирует фермент протеинкиназу А, которая в свою очередь фосфорилирует бифункциональный фермент- 2,6-фруктозобифосфатазу/6-фосфофрукто-2-киназу. Фосфорилированный фермент утрачивает КИНАЗНУЮ активность (превращение фруктозо-6-фосфата во фруктозо-2,6-дифосфат), сохраняя при этом фосфатазную активность(превращение фруктозо-2,6-дифосфата во фруктозо-6-фосфат).
 Фруктозо-2,6-дифосфат
(Ф-2,6-ДФ) является
активатором 6-фосфофрукто-1-киназы и
ингибитором фруктозо-1,6-бифосфатазы,
поэтому снижение его количества в
результате воздействия глюкагона
приводит к подавлению гликолиза и
активации процесса глюконеогенеза.
Такой механизм действия характерен
только для клеток печени, где имеется
одна из изоформ бифункциональной
фосфатазы/киназы
(белок из 470 аминокислотных остатков).
Фруктозо-2,6-дифосфат
(Ф-2,6-ДФ) является
активатором 6-фосфофрукто-1-киназы и
ингибитором фруктозо-1,6-бифосфатазы,
поэтому снижение его количества в
результате воздействия глюкагона
приводит к подавлению гликолиза и
активации процесса глюконеогенеза.
Такой механизм действия характерен
только для клеток печени, где имеется
одна из изоформ бифункциональной
фосфатазы/киназы
(белок из 470 аминокислотных остатков).
 В
клетках сердечной мышцы имеется другая
изоформа (530 АК), с ПРОТИВОПОЛОЖНЫМ
влиянием глюкагона. Механизм ускорения
инсулином процесса гликоза до конца не
выяснен, имеется предположение, что в
нем принимают участие некоторые,
неизвестные пока посредники, активирующие
протеинфосфатазу, результатом действия
которой является ДЕФОСФОРИЛИРОВАНИЕ
бифункциональной фосфофруктофосфатазы/киназы,
что увеличивает ее киназную активность
(см.выше) и снижает фосфатазную. Следствием
является накопление фруктозо-2,6-дифосфата
и ускорение гликолиза.
В
клетках сердечной мышцы имеется другая
изоформа (530 АК), с ПРОТИВОПОЛОЖНЫМ
влиянием глюкагона. Механизм ускорения
инсулином процесса гликоза до конца не
выяснен, имеется предположение, что в
нем принимают участие некоторые,
неизвестные пока посредники, активирующие
протеинфосфатазу, результатом действия
которой является ДЕФОСФОРИЛИРОВАНИЕ
бифункциональной фосфофруктофосфатазы/киназы,
что увеличивает ее киназную активность
(см.выше) и снижает фосфатазную. Следствием
является накопление фруктозо-2,6-дифосфата
и ускорение гликолиза.
БРОЖЕНИЕ- еще одна разновидность АНАЭРОБНОГО расщепления глюкозы (нехарактерно для человеческого организма):
С6Н12О6 2СО2 + 2 С2Н5ОН
Другой путь- молочнокислое брожение:
С6Н12О6 2 СН3СН(ОН)СООН
ВТОРОЙ ЭТАП-окислительное декарбоксилирование с образованием ацетил-КоА:
СН3С(=О)СООН + КоА-SH + NAD+ СН3С(=О)-S-КоА + NADH + CO2 Окисление ацетил-КоА до СО2 и Н2О протекает на третьем этапе окисления глюкозы- в цикле трикарбоновых кислот.
Произошедшие в цитоплазме реакции гликолиза приводят к образованию пирувата (ПВК). Последующие превращения пирувата протекают во внутримитохондриальном пространстве- матриксе митохондрии. В матрикс молекулы ПВК приникают через внутреннюю митохондриальнубю мембрану в результате обмена с ионами гидроксила (НО-). Уже в матриксе, при участии NAD-зависимой пируватдекарбоксилазы ПВК превращается в ацетил-КоА:

Образующийся NADH генерирует в дыхательной цепи 6 молекул АТФ (в пересчете на 1 молекулу глюкозы).
Ацетил-КоА вступает в цикл трикарбоновых кислот (ЦТК, цитратный цикл, цикл Кребса) при взаимодействии с молекулой оксалоацетата:

Реакция катализируется цитрат-синтазой. Другие реакции цикла и катализаторы:

Энергетический баланс окисления ацетил-КоА в ЦТК составляет 24 молекулы АТФ. Полное окисление глюкозы (гликолиз (8 АТФ)+ ЦТК (24 АТФ), включая окислительное декарбоксилирование пирувата в цетил-КоА (6 АТФ)), дает 38 молекул АТФ на молекулу глюкозы.
Реакции цикла превращений Ацетил-КоА впервые были установлены Гансом Кребсом (университет Шеффилда, Великобритания), за что в 1953 году он был удостоен Нобелевской премии.
Активность ЦТК повышается с ростом концентрации АДФ (аллостерический активатор изоцитрат-дегидрогеназы) и снижается с ростом АТФ. За один оборот в ЦТК молекула ПВК окисляется полностью в 3 Н2О и СО2.
ГЛИОКСИЛАТНЫЙ ЦИКЛ
Запас промежуточных продуктов ЦТК
пополняется специальными ферментативными
механизмами. Такова, в частности, функция
глиоксилатного цикла. Глиоксилатный
цикл (ГЦ)- видоизмененный ЦТК. В нем
ферменты изоцитратдегидрогеназа
и a-кетоглутаратдегидрогеназа
заменены ферментами изоцитрат-лиазой
и малат-синтазой.
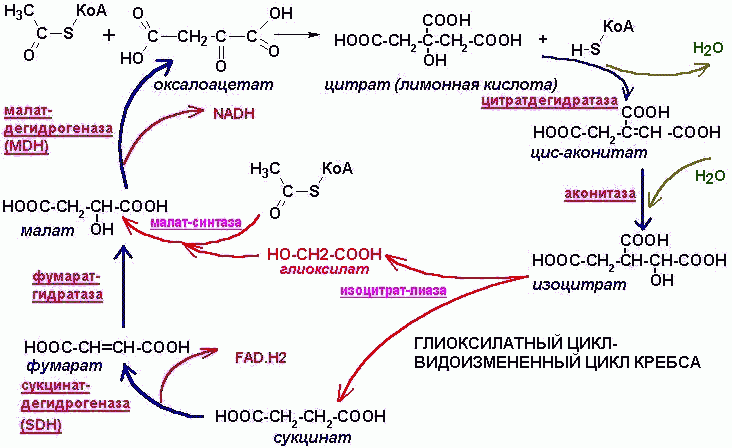 Первая
расщепляет лимонную кислоту до янтарной
и глиоксиловой (СООН-СН2-СН2-СООН и
СООН-СНО). Малат-синтаза синтезирует из
ацетил-КоА и глиоксиловой кислоты
яблочную кислоту:
Первая
расщепляет лимонную кислоту до янтарной
и глиоксиловой (СООН-СН2-СН2-СООН и
СООН-СНО). Малат-синтаза синтезирует из
ацетил-КоА и глиоксиловой кислоты
яблочную кислоту:
СН3-СО-S-КоА + Н2О + СООН-СНО ® СООН-СН(ОН)-СН2-СООН + КоА-SH
Таким образом, при каждом обороте цикла в него вкладывается 2 молекулы ацетил-КоА, образуется одна молекула янтарной кислоты, которая может вступать в процессы глюконеогенеза, ЦТК и др. процессы. Туда же могут идти яблочная и лимонная кислоты.
ФОСФОГЛЮКОНАТНЫЙ ПУТЬ
(пентозофосфатный цикл)
Кроме гликолиза в клетках существует еще один путь расщепления углеводов- фосфоглюконатный путь (ФГП). Посредством ФГП одна молекула глюкозы расщепляется на СО2 и рибозо-5-фосфат. Превращения фосфоглюконатного пути многочисленны и сложны, однако суммарный результат его превращений описывается довольно простым уравнением:
6-Фосфоглюконолактон + Н2О + NADP+ ® Рибозо-5-Ф + СО2 + NADPН + Н+

БИОЛОГИЧЕСКАЯ ФУНКЦИЯ ФОСФОГЛЮКОНАТНОГО ПУТИ
1) АМФИБОЛИЧЕСКАЯ (распад углеводов и синтез NADPH и рибозо-5-фосфата)
2) ЭНЕРГЕТИЧЕСКАЯ (подключение его продукта- глицеральдегид-3-фосфата к гликолизу дает выход энергии)
Полученные в различных метаболических циклах восстановительные эквиваленты (NADH, NADPH, FAD.H2) утилизируются в дыхательной цепи, где при их окислении синтезируется АТФ (сопряженное или окислительное фосфорилирование), либо используются в других метаболических путях при синтезе макромолекул.