

От нуклеотидов к основаниям.
Гуаниновые нуклеотиды гидролизуются с образованием гуанозина, который подвергается фосфоролизу до гуанина и рибоза 1-Ф. Гуанин дезаминируется гуанин дезаминазой с образованием ксантина. Аденозин также можно получить по такому пути, однако внутриклеточные нуклеотидазы у человека не очень активны по отношению к AMФ. АМФ чаще дезаминируется ферментом аденилат (AMФ) дезаминазой с образованием ИМФ. Последний далее гидролизуется нуклеотидазой с образованием инозина и после фосфоролиза превращается в гипоксантин.
Н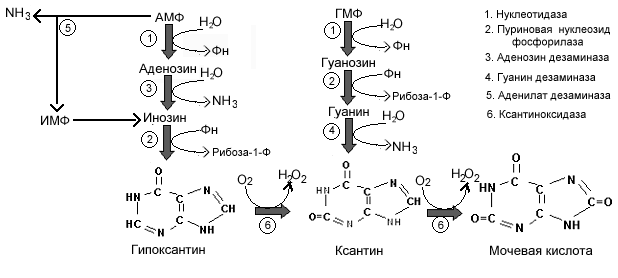 екоторое
количество аденозина образуется изS-аденозилметионина в
процессах переметилирования. Аденозин
дезаминируется до инозина аденозин
дезаминазой. Недостаточность аденозин
дезаминазы или пуриновой нуклеозид
фосфорилазы ведет к двум различным
болезням иммунодефицита механизмами,
которые до конца не раскрыты.
екоторое
количество аденозина образуется изS-аденозилметионина в
процессах переметилирования. Аденозин
дезаминируется до инозина аденозин
дезаминазой. Недостаточность аденозин
дезаминазы или пуриновой нуклеозид
фосфорилазы ведет к двум различным
болезням иммунодефицита механизмами,
которые до конца не раскрыты.
Рис 8-10. Основные пути катаболизма нуклеотидов
При недостаточности аденозин дезаминазы, страдают TиB-лимфоциты, а при недостаточности фосфорилазы нарушается функцияTклеток, аBклетки остаются нормальными. В сентябре 1990 г была успешно применена генинженерная технология для лечения 4-летней девочки с недостаточностью аденозин дезаминазы.
Катаболизм метилированных (минорных) пуринов зависит от расположения метильной группы. Если метильная группа связана с группой -NH2, она удаляется вместе с -NH2, и оставшаяся часть обменивается в дальнейшем обычным способом. Если метил связан с атомом азота гетероцикла, соединение выделяется в неизменном виде с мочой.
От оснований к мочевой кислоте
И адениновые и гуаниловые нуклеотиды превращаются в одно общее промежуточное соединение ксантин. Гипоксантин, возникающий из аденина, окисляется в ксантин ксантиноксидазой . Гуанин дезаминируется с образованием аммиака и ксантина. Образующийся аммиак переносится к печени в составе глутамина, где используется для синтеза мочевины.
Ксантин, подобно гипоксантину, окисляется кислородом и ксантиноксидазой в мочевую кислоту с образованием перекиси водорода. У человека, мочевая кислота выделяется, а перекись водорода разрушается каталазой. Высокая активность ксантиноксидазы обнаруживается только в клетках печени и кишечника.
Р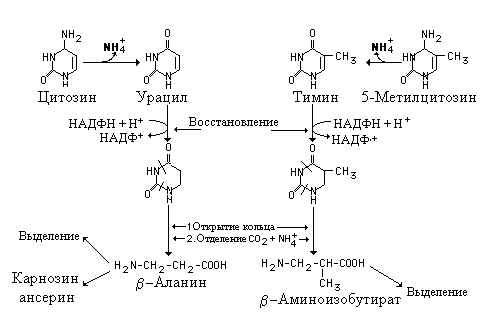 ис
8.11Основные
пути катаболизма пиримидиновых азотистых
оснований
ис
8.11Основные
пути катаболизма пиримидиновых азотистых
оснований
Катаболизм пиримидиновых нуклеотидов
В отличие от пуринов, кольцевая структура пиримидинов разрушается с образованием обычных конечных продуктов катаболизма - -аминокислот, аммиака и двуокиси углерода. В катаболизме пиримидиновых нуклеотидов принимают участие нуклеотидазы и пиримидиновые нуклеотид фосфорилазы, которые превращают мононуклеотиды в свобоные основания. Аминогруппы цитозина и 5-метилцитозина отделяется в форме аммиака..
Дециклизация. Свободные пиримидиновые основания восстанавливаются НАДФН+Н+. АтомыN2 иC3 пиримидинового кольца выделяются в форме аммиака и двуокиси углерода соответственно. Оставшаяся часть кольца представляет собой-аминокислоту .-аминоизомасляная кислота образуется из тимина или 5-метилцитозина и в основном затем выделяется почками. Незначительная часть ее после переаминирования может превращаться в сукцинил-КоА и использоваться в цикле трикарбоновых кислот. Из цитозина и урацила образуется-аланин, который может быть использован для синтеза биологически активных дипептидов карнозина (гис--ала) или ансерина (метил гис--ала) в мозге и мышцах.