

Некоторые перестройки генетического материала могут быть восстановлены.
![]()
 Репаративные
механизмы, которые использует клетка
для поддержания стабильности информации,
заложенной в ДНК универсальны -
функциональная, а иногда и структурная
гомология элементов, образующих эти
механизмы, прослеживается от бактерий
до человека.
Репаративные
механизмы, которые использует клетка
для поддержания стабильности информации,
заложенной в ДНК универсальны -
функциональная, а иногда и структурная
гомология элементов, образующих эти
механизмы, прослеживается от бактерий
до человека.
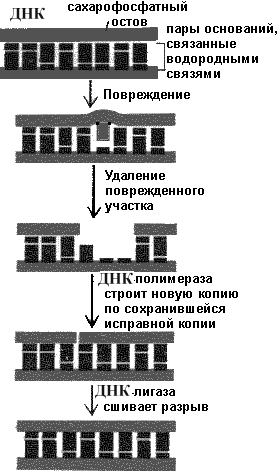
Рис.8.29. Общие принципы репаративного синтеза ДНК.
Чем сложнее клетка, тем большее количество структурных и регуляторных генов и их продуктов участвуют в процессах репарации ДНК, хотя принципиальная схема конкретного процесса, как правило, остается неизменной.
Ключевой частью всех репаративных систем является распознавание дефекта в структуре ДНК, который затем или сразу восстанавливается или маркируется специфическими белками.
Эксцизионная репарация ведущий способ восстановления изменений структуры ДНК
Основной механизм репарации большинства дефектов получил название эксцизионной репарации, в основе которой лежит удаление измененного основания или нуклеотида. Различают два принципа такой репарации - эксцизионная репарация оснований (ЭРО) и эксцизионная репарация нуклеотидов (ЭРН). Биологическая функция ЭРО заключается в восстановлении исходной структуры модифицированных оснований ДНК. Эта система высокоспецифична и состоит из специализированных ДНК-N-гликозилаз , которые опознают и удаляют поврежденные основания, АР-эндонуклеаз или АР-лиаз , которые, соответственно, расщепляют нить ДНК с 5'- или 3'-конца апуринового или апиримидинового (АР) участка; фосфодиэстераз (соответственно 5'или 3'), которые выщепляют дезоксирибофосфатный остаток, и, наконец, ДНК-полимеразы и ДНК-лигазы . К концу 90-х г.г. из бактерий (E. coli, Micrococcus luteus и др.) выделено до 8 различных N-гликозилаз, способных репарировать все известные модификации оснований. Несколько различных Nгликозилаз обнаружены и в клетках человека. В целом, система ЭРО - чрезвычайно действенный барьер мутациям оснований. Обычно цепь реакций ЭРО начинается с обнаружения и удаления одного из оснований, катализируемого соответствующим ферментом группы AP-эндонуклеаз типа II (апуриновых / апиримидиновых эндонуклеаз). Затем происходит гидролитическое расщепление 3'-фосфоэфирной связи, причем фосфат остается в 5'-положении уходящей цепи ДНК. Далее с 3'-концом в месте расщепления цепи связывается-ДНК-полимераза , катализирующая гидролитическое удаление дезоксирибозилфосфатного остатка, таким образом освобождая 3'-конец для включения соответствующего канонической паре нуклеотида. Далее, после включения в освободившееся положение соответствующего нуклеотидного остатка, ДНК-лигаза воссоединяет непрерывную цепь ДНК. Как в любой другой репаративной реакции, в системе ЭРО завершающим является ресинтетический этап, восстанавливающий правильность спаривания с помощью ДНК-полимеразы I (у E. coli) и ДНК-лигазы. Генетических заболеваний, сопряженных с нарушением системы ЭРО пока не обнаружено.
В отличие от реакций ЭРО, которые специфичны для достаточно узкого круга повреждений ДНК, система ЭРН, хотя и с разной эффективностью, удаляет все возможные повреждения, и потому роль ее в поддержании стабильности генома велика. ЭРН детально изучена у E. coli и активно изучается в клетках дрожжей и человека, причем у человека благодаря раскрытию генетической природы таких заболеваний, как пигментная ксеродерма (XP), синдром Кокейна ( СS ) и трихотиодистрофия ( ТТD ) Стратегия ЭРН во всех случаях, от бактерии до человека одинакова, хотя по мере усложнения объекта количество задействованных в реакции генов и их продуктов возрастает с 6-8 у E. coli до 30 у человека. Основные этапы ЭРН подобны ЭРО.
1) узнавание повреждения и изменения конформации ДНК вокруг него;
2) раскручивание участка ДНК в области повреждения;
3) эксцизия поврежденного участка ДНК и
4) ресинтетический этап, осуществляемый ДНК-полимеразой и ДНК-лигазой
Хотя детали процесса ЭРН у человека не ясны, так как не определена роль целого ряда генов, в первом приближении он выглядит так. Белок ХРА в комплексе с белком связывающим одноцепочечные ДНК RPA ( репликационным белком А ), перемещаясь вдоль одноцепочечной ДНК, опознает конформационнoе повреждениe. Взаимодействуя через другой участок белка ХРА с базальным фактором транскрипции TFIIH (две из субъединиц которого, белки ХРВ и ХРD , обладают геликазной активностью с противоположной ориентацией раскручивания двухцепочечной ДНК), они образуют комплекс, который расплетает ДНК в области повреждения (в состав этого комплекса входит и белок ХРС с неясными функциями). Через третий участок белка ХРА к комплексу примыкает гетеродимер ERCC1-XPF , который вносит однонитевой разрыв в ДНК с 5'-конца на расстоянии 16-25 нуклеотидных пар. от повреждения, тогда как белок XPG , входящий в комплекс белков эксцинуклеазы через взаимодействие с белком RPA, делает надрез с 3'-конца на расстоянии 2-9 нуклеотидных пар. (в разрезании принимает участие и белок ХРС, а белок ХРЕ активирует реакцию). В результате бимодальной инцизии участок ДНК размером около 29 нуклеотидов. высвобождается, а образующаяся брешь ресинтезируется с помощью ДНК-полимеразы илиДНК-полимеразы, сопутствующего репликации фактора PCNА, репликационного фактора C-RFС и ДНК-лигазы I . Представленная картина во многом основана на реконструировании процесса в открытой системе с участием 10 хорошо очищенных белков и не охватывает всех возможных участников этого процесса.