

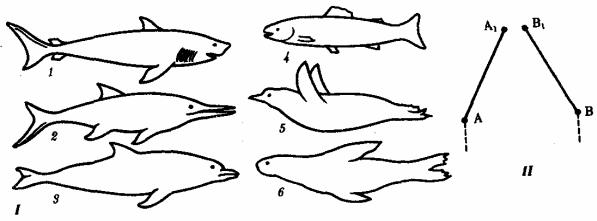
экологические задачи они решают сходным образом. Конвергентные адаптации возникают в этом случае на разной генетической основе, затрагивают в первую очередь поверхностные признаки, не распространяясь на общий план строения и наиболее существенные черты организации соответствующих групп. Примером конвергентной эволюции являются форма тела и особенности локомоции в воде у акуловых рыб, водных пресмыкающихся — ихтиозавров, костистых рыб, пингвинов, ластоногих и китообразных млекопитающих, внутреннее строение которых полностью соответствует особенностям, характерным для классов, к которым они относятся (рис. 13.6).
Другая форма соотносительной эволюции — параллелизм — реализуется в двух или нескольких группах, связанных более или менее отдаленным родством, которое основано на дивергенции от общего предка. В связи с общностью части генофондов, унаследованных от предков, у них возникают сходные адаптации в условиях действия факторов отбора в одинаковом направлении. Параллельное филетическое развитие двух родственных групп обеспечивается реализацией закона гомологических рядов (см. разд. 13.3.5). Примером параллелизма является эволюция одногорбого и двугорбого верблюдов соответственно в Африке и Центральной Азии, имеющих сходные адаптации к жизни в пустынях
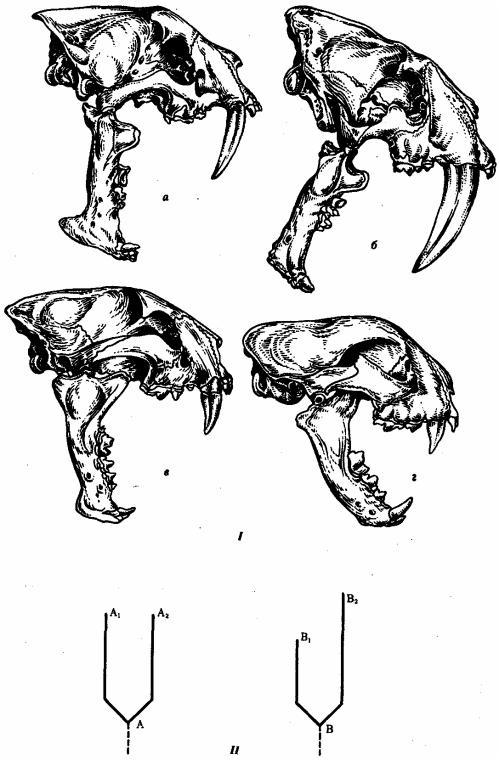
Параллелизм можно наблюдать и в родственных группах организмов, разобщенных временем. Так, во второй половине кайнозойской эры в семействе кошачьих такой признак, как саблезубость (гипертрофия клыков верхней челюсти), возникал по крайней мере четырежды у представителей четырех разных родов (рис. 13.7).
13.1.4. Биологический прогресс и биологический регресс
Эволюция любого типа — арогенная, аллогенная или идущая по типу мофофизиологического регресса в природе — в целом приводит к всесветному расселению живых организмов разного уровня организации.
57
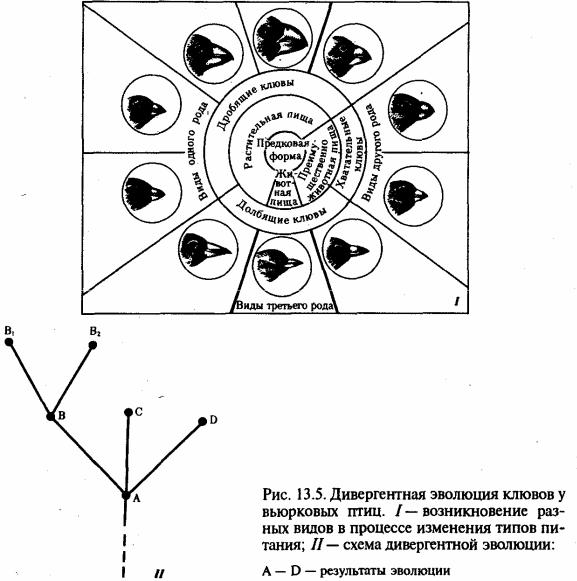
Рассматривая эволюцию отдельных таксонов, можно убедиться в том, что некоторые из них находятся в состоянии расцвета, в то время как другие вымирают. Успех группы организмов в эволюционном процессе оценивают как состояние биологического прогресса, критериями которого являются: 1) увеличение количества представителей соответствующей группы; 2) расширение ареала распространения;
3) активизация видоообразования в роде, увеличение количества родов в семействе, семейств в отряде и т.д.
Явление, противоположное биологическому прогрессу, — биологический регресс — свидетельствует об угасании филогенетической группы, ведущем к ее вымиранию. Так же как и биологический прогресс, состояние регресса может длиться очень долго. Оставшиеся немногочисленные представители некогда биологически прогрессивной группы, найдя соответствующую их организации экологическую нишу, могут продолжать существовать на протяжении сотен миллионов лет. Они называются реликтами, обладают консервативностью
58
организации и крайне медленно эволюционируют. Примерами реликтов являются современная кистеперая рыба — латимерия, ближайшие родственники которой жили в девонском периоде более 200 млн. лет назад, а также дерево гинкго, непосредственные предки которого существовали на Земле уже в первой половине мезозойской эры.
Рис. 13.6. Конвергентная эволюция формы тела позвоночных. I—водные животные разного происхождения; II—схема конвергентной эволюции:
1—акула, 2—ихтиозавр, 3—дельфин, 4—костистая рыба, 5—пингвин, 6— тюлень; A—A1, B—B1 — этапы эволюции
На протяжении исторического развития одна и та же группа может претерпевать периоды бурного расцвета, стабильного состояния, или стагнации, и упадка. Так, в девонском периоде появились первые земноводные, вставшие на путь арогенной эволюции. Заселив наземную среду обитания, они господствовали на ней в течение 75 млн. лет. На границе палеозойской и мезозойской эр (230 млн. лет назад) бурный биологический прогресс претерпевают пресмыкающиеся, быстро вытесняющие земноводных, которые с тех времен и до современного состояния переживают биологический регресс.
Расцвет пресмыкающихся длился до последней трети мезозойской эры, когда (около 65 млн. лет назад) он сменился биологическим регрессом, после чего освобождающиеся вымирающими динозаврами экологические ниши стали занимать бурно эволюционирующие млекопитающие и птицы, биологический прогресс которых продолжается до настоящего времени. В то же время такой класс, как костистые рыбы, пройдя период бурного расцвета к началу мезозойской эры и прочно заняв разнообразные места обитания, до настоящего времени находится в состоянии стабильного господства среди позвоночных в водной среде обитания.
Состояние биологического прогресса может быть обусловлено как аро- и аллогенезом, так и морфофизиологическим регрессом. Действительно, занятие новых экологических ниш как в новых адаптивных зонах, так и в уже освоенных
59
приводит и к активизиции размножения, и к расширению ареала, и к усилению видообразования.
Рис. 13.7. Параллельная эволюция саблезубости у кошачьих. I—черепа саблезубых кошек в разные периоды кайнозойской эры; II—схемы параллельной эволюции:
а—махайрод (олигоцен - 37-25 млн. лет назад), б—смилодон (миоцен - 25-9 млн. лет
60