

20 . Тканевое дыхание.
Тканевое дыхание– один из процессов диссимиляции, по сути это есть биологическое окисление в тканях и клетках организма. В организме существует три пути потребления и утилизации кислорода:
1 – й путь: 90 – 95 % кислорода идёт на митохондриальное окисление.
2 – й путь: 5 – 10 % идёт на микросомальное окисление (в печени при поступлении токсинов).
3 – й путь: 2 – 5 % - перекисное окисление.
21 . Дыхательная цепь.
В процессе окисления ацетилКоА в цикле Кребса восстановленные формы НАД * Н2и ФАД * Н2 поступают в дыхательную цепь, где энергия электрона и протона трансформируется в энергию макроэргических связей АТФ.Дыхательная цепь– это совокупность дегидрогеназ, которые транспортируют электроны и протон с субстрата на кислород. Принцип функционирования дыхательной цепи основан на 1 – ом и 2 – ом законах термодинамики (1 – й закон: закон сохранения энергии; 2 – й закон: все системы стремятся к минимальной энергии).
Дыхательная цепь локализуется во внутренней мембране митохондрий и имеет два пути введения электрона и протона или два входа; дегидрогеназная цепь образует четыре комплекса:
1 – й вход – НАД - зависимый (поступают электрон и протон со всех НАД - зависимых реакций)
2 – й вход – ФАД - зависимый (аналогично)
Кофермент Q или убихинон – это гидрофобное соединение, является компонентом клеточной мембраны (находится на наружной поверхности), содержится в большой концентрации, относится к группе витаминов.
О
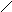
 //
//
Н3СО СН3 СН3СН3



 /
/
/
/



 Н3СО
(СН2
– СН = С – СН2)9
– СН2– СН = С – СН2
Н3СО
(СН2
– СН = С – СН2)9
– СН2– СН = С – СН2
\\
О
КОФЕРМЕНТ Q .
ЦИТОХРОМ.
СН2
//
СН СН3
/ /













 Н3С
СН = СН2
Н3С
СН = СН2



 NN
NN






 Fe
Fe







 NN
NN

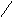

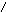
 Н3С
СН3
Н3С
СН3





\ \
СН2 СН2– СН2 – СО2
\
СН2
\
СО2
ЦИТОХРОМ aa3 – (цитохромоксидаза)
CH3 CH3 CH3
| | |
СН2 – СН – (СН2 )3– СН –(СН2)3–CH
| |
HO- СНCH3 CH3


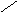


 | |
| |



 H3C-
- CH = CH2
H3C-
- CH = CH2



 N N
N N



 Fe
Fe





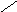







 N N
N N
O = C -
|
H | |
CH2 CH2 – CH2 – CO2
| |
CH2
|
CO2
Цитохромоксидаза имеет высокую степень сродства к О2 и может работать при его низких концентрациях.аа3 состоит из шести субъедениц, каждая из которых содержит гем и атомCu. Две субъеденицы составляют цитохрома, а остальные четыре относятся к цитохромуа3.
Дыхательная цепь активно реагирует на чрезмерные нагрузки, яды. Так, например, барбитураты ингибируют перенос электрона и протона в первый комплекс дыхательной цепи, в ответ на это возникает энергетический голод ткани. Это происходит потому, что возникает недогрузка дыхательной цепи протонами и электронами. В результате снижается синтез АТФ и в ткани активируется производство и окисление эндогенной янтарной кислоты. Такое же действие оказывает и алкоголь, так как этанол является НАД – зависимым субстратом. Этанол монополизирует основной фонд НАД и превращает его в НАД * Н2. Дыхательная цепь перегружается НАД * Н2и ткань не получает энергии за счёт окисления естественных субстратов. Такие яды, как цианиды, СО, Н2S, блокируют четвёртый комплекс дыхательной цепи. Если ингибирование первого комплекса приводит к активации трёх оставшихся и за счёт этого дыхательная цепь компенсируется, то при блокировании четвёртого комплекса работа дыхательной цепи не состоится, что приводит к смерти. Четыре комплекса, входящие в состав дыхательной цепи, неравноценны: в 1,3 и 4 происходит синтез АТФ (АДФ + Фн).
Работа всей дыхательной цепи состоит из двух составляющих: 1) образование АТФ (50% энергии депонируется в виде АТФ); 2) рассеивание остальных 50 % энергии в виде тепла.
Для оценки эффективности работы дыхательной цепи и окислительного фосфорилирования в 1939 году введён коэффициент Р / О–показатель степени спряжения фосфорилирования и дыхания. Р / О – отношение количества поглощённых молекул фосфата (в процессе АДФ + Фн) к количеству поглощённого кислорода. Так, если окисляются НАД – зависимые субстраты, то электроны в этом случае проходят все три стадии фосфорилирования и на образование АТФ уходят три молекулы Фни один атом О2, поэтому Р / О = 3. Если окисляются ФАД – зависимые субстраты, то электроны проходят только две стадии фосфорилирования и тратится две молекулы Фнна атом О2, поэтому Р / О = 2. При окислении витамина С и адреналина, электроны поставляются прямо на цитохромс, поэтому они проходят только одну стадию фосфорилирования и поэтому Р / О = 1.
В ряде случаев некоторые пункты фосфорилирования могут «выключаться» - такое состояние называется разобщением окислительного фосфорилирования. В этом случае Р / О снижается: для
НАД – зависимых субстратов – ниже трёх, для ФАД – зависимых субстратов – ниже двух.
Исходя из первого закона термодинамики, в разобщённых митохондриях увеличивается теплообразование. Это происходит за счёт того, что та энергия электронов, которая должна была быть использована для синтеза АТФ в «выключенной» стадии фосфорилирования, рассеивается в виде тепла. Процесс разобщения окислительного фосфорилирования лежит в основе лихорадки, вызванной бактериями, вирусами и другими агентами. Разобщение резко увеличивается при охлаждении организма. Работамитохондрий при всех «выключенных» стадиях фосфорилирования называетсясопряжённой, в противном, выше описанном случае,разобщённойидыханиепри этом называетсясвободным. В качестверазобщителей окислительного фосфорилирования выступают слабые гидрофобные кислоты (жирные кислоты), тироидные гормоны, лекарства.
Дыхательная цепь имеет механизм шунтирования: сброс электрона и протона с НАД на цитохромы или с НАД на межмембранные дегидрогеназы, не нарушая мембрану и гладкую эндоплазматическую сеть. Такой перенос характерен для печени. При воздействии какого – либо блокатора, возникает блок в первом комплексе дыхательной цепи. Происходит накопление НАД * Н2, и становится реальной угроза гипоксии и печень может погибнуть. Чтобы этого не произошло, происходит сброс НАД * Н2с митохондриальной дыхательной цепи на микросомальную.