BKh_Kolokvium_2_uglevody
.doc|
1. Переваривание и всасывание углеводов в ЖКТ. Мальабсорбция. Причины, клинические проявления.Переваривание углеводов начинается в ротовой полости. Под действием ферментов слюны а-амилазы. Крахмал расщепляется на декстрины и на мальтозу(если долго нах-ся У в ротовой полости.Амилаза слюны активирует Са2+ и Сl-. В желудке нет ферментов т.к. рН = 1,5-2,5, следов. переваривания нет. Главное место переваривания У – 12пк под дуйствием амилазы поджелудочной железы (крахмал,гликоген->мальтоза).Мальтоза распадается на 2 молекулы глюкозы под действием мальтазы. Продуктом полного переваривания крахмала и гликогена является глюкоза. Различают три вида пищеварения : 1) полостное (неэффективно , так как вероятность встречи фермента и субстрата не велика и подчиняется закону Брауновского движения ) 2) пристеночное (осуществляется в гликокаликсе , который представляет собой гликопротеиновый комплекс , локализованный над и под микроворсинками тонкого кишечника) 3) внутриклеточное ( проходит по механизму фаго и пиноцитоза ; является несовершенным, поскольку может приводить к развитию аллергических реакций. Всасывания осуществляются тремя путями : 1) пассивная диффузия (по градиенту концентрации) , так переносятся манноза , арабиноза , ксилоза 2) облегчённая диффузия (облегчается путём образования гидрофобных каналов и пор при контакте мембраны с транспортируемым веществом ) 3) активный транспорт ( против градиента концентрации , за счёт энергии макроэргических связей АТФ или энергии мембранного потенциала )
Мальабсорбция (нарушение всасывания в кишечнике) – это синдром, характеризующийся набором клинический проявлений, развивающихся вследствие нарушения пищеварительной и транспортной функций тонкого кишечника, что в свою очередь ведет к патологическим изменениям обмена веществ. Клинические проявления: диарея, стеаторея, вздутие и урчание, иногда болезненность в животе, слабость, утомляемость, апатия. Причины: нарушением водно-электролитного гомеостаза и недостаточностью необходимых для нервной системы веществ; недостаточностью витаминов и микроэлементов
|
2. Механизм транспорта глюкозычерез мембраны. Различают транспорт: клеточный и пераклеточный (имеет второстепенное значение) Механизм выглядит так : глюкоза соединяется с ионами Na , образуя комплексное соединение , которое транспортируется внутрь клетки. Затем комплекс распадается, а освобождёный ион Na транспортируется обратно. Попавшая внутрь клетки глюкоза подвергается фосфорилированию. фосфорилирования глюкозы: Гл 0 -> Г6Ф2- Причинами этого является: 1) при фосфорилировании глюкоза приобретает заряд, это позволяет ей эффективно взаимодействовать с активным центром ферментов, катализирующих последующие реакции. 2) образовавшийся глюкоза – 6 – фосфат имеет отрицательный заряд, что препядствуют его выходу из клетки, то есть срабатывает эффект « замерзания» 3) фосфат, который связывается с глюкозой становится макроэргическим.
|
3. Схема обмена Г-6-Ф в организме. Гормональная регуляция обмена глюкозы. Гл
GLUT
ПФП Г-6-Ф глюкоген
ГАГ ПВК лактат 2Н2
ацетил КоА ЦТК БО
СО2+Н2О АТФ
Все метаболические пути глюкозы находятся под влиянием инсулина , то есть инсулинзависимы. Инсулин повышает проницаемость тканей для глюкозы. Все ткани, кроме печени и головного мозга, инсулинзависимы , следовательно концентрация глюкозы в этих органах будет зависить от концентрации глюкозы в крови. При извлечении инсулина все 5 путей метаболизма будут нарушены.
|
4. Строение гликогена. Синтез и фосфоролиз гликогена.
Гликоген – большая ветвистая молекула с молекулярной массой 106-107 Дальтон. Линейные участки молекулы гликогена связаны α(1→4) связью, точки ветвления представлены α(1→6) гликозидной связью. Синтез: Глюкоза+АТФГ-6-Ф + АДФ (гексогеназа) Г-1-Ф УДФ-глюкоза (глюкоген)n+1 Глюкогенолиз - активируется стрессом. (гормоны-адренолин и глюкагон) Фосфоролиз: (глюкоген)n+1(глюкоген)n-1+Г-1-Ф (гликогенфосфорилаза) Г-6-Ф (фосфоглюкомутаза) Глюкоза + H3PO4 (фосфатаза глюкозо-6 фосфат) |
5.Гормональная регуляция метаболизма гликогена. Роль цАМФ, ионов Са в регуляции гликогенеза и гликогенолиза. В регуляции синтеза гликогена ключевую роль играет гликогенсинтаза. Фермент находится в клетке в неактивном, фосфорилированном, состоянии и называется гликогенсинтаза D, т.е. активность его зависит от глюкозо-6-фосфата (аллостерический активатор) и гормона инсулина. Инсулин активирует фермент фосфатазу, который и превращает гликогенсинтазу D в активную форму – гликогенсинтазу I . Адреналин и глюкагон, активируя аденилатциклазу, способствуют образованию цАМФ, который запускает «каскадный механизм» фосфорилирования ферментов распада и синтеза гликогена. В результате фосфорилирования образуется активная, гликогенфосфорилаза и неактивная гликогенсинтаза. В этих условиях будет осуществляться распад гликогена. Напротив, под действием инсулина, включающего механизм дефосфорилирования ключевых ферментов, появятся неактивная гликогенфосфорилаза, и активная, гликогенсинтаза. В этих условиях будет происходить синтез гликогена. Активность обоих обоих ключевых ферментов синтеза и распада гликогена в печени регулируется также Са++ – выход кальция из внутриклеточных депо в цитозоль клетки контролируется инозитолтрифосфатом (ИТФ), который освобождается фосфолипазой С при распаде входящего в состав мембраны фосфатидилинозитол пирофосфата. |
|
|
6. Особенности глюкогенолиза в печени и мышцах. Регуляция активности ферментов. В печени находится Глюкозо-6-фосфатаза – важнейший фермент, ответственный за образование глюкозы. Печень является основными поставщиками глюкозы для тканей организма. Глюкозо-6-фосфатаза практически отсутствует в мышцах – миоциты получают глюкозу из крови. Кроме того, в мышцах синтезируется глюкозо-6-фосфат из глюкозо-1-фосфата в процессе распада гликогена. Между гликолизом, протекающим в мышцах при их интенсивной работе, и глюконеогенезом, осуществляемым печенью, существует тесная взаимосвязь (цикл Кори): образующая в мышцах молочная кислота поступает в общий кровоток, захватывается печенью и используется ею в качестве субстрата глюконеогенеза; синтезируемая при этом глюкоза отдаётся в кровототок и метаболизируется мышцами для получения энергии.
Регуляция осуществляется путём аллостерического контроля ферментов, которые для этих двух путей различны: 1. Активности гексокиназы и глюкозо-6-фосфатазы регулируются уровнем глюкозо-6-фосфата: гексокиназа им ингибируется, а фермент глюконеогенеза активируется. 2.Для пируваткиназы и пируваткарбоксилазы аллостерческим эффектором является ацетил-КоА, однако если для фермента глюконеогенеза он является положительным модулятором, то активность пируваткиназы ацетил-КоА, напротив, ингибирует. 3. Главным аллостерическим регулятором двух взаимосвязанных путей – гликолиза и глюконеогенеза – является фруктозо-2,6-дифосфат: увеличение его концентрации активирует ключевой фермент гликолиза – фосфофруктокиназу-1; снижение его концентрации активирует конкурирующую реакцию – образование фруктозо-6-фосфата, т.е. приводит к усилению глюконеогенеза. |
7. Кинетическое различие в действии глюкокиназы и гексокиназы. Гексагеназа: *высокое сродство к глюкозе Кm > 0.1нм *высокая специфичность *локализация: печень, поджелудочная железа. * регулирует процесс гликолиза
Гексакиназа: *низкое сродство к глюкозе Km= 10нм *низкая спецефичность *не ингибирует Г6Ф и относ АТФ/АДФ
|
8. Метаболизм галактозыи фруктозы в норме и при патологии Норма (фруктоза): фруктокиназа фосфофруктокиназа фруктоза+АТФ------------Ф-1-Ф+АТФ ------------Ф-1,6-Ф
г альдегид
Гликолиз ГНГ глицерин Нарушения метаболизма фруктозы Недостаточность фруктокиназы клинически не проявляется. Фруктоза накапливается в крови и выделяется с мочой, где её можно обнаружить лабораторными методами. галактокиназа
Гл-6-Ф
Гликоген клюкоза гликолиз крови Галактоземия, вызвана недостаточностью галактозо-1-фосфатуридилтрансферазы (ГАЛТ). Это заболевание проявляется очень рано, и особенно опасно для детей, так как основным источником углеводов для них служит материнское молоко, содержащее лактозу.
|
9. Гликолиз. Реакции, классиф., регуляция и биолог.роль. Энергетич баланс фнаэробного гликолиза. Регуляция гликолиза. Гликолиз – процесс анаэробного распада глюкозы. Происходит в цитоплазме клетки. В анаэробных условиях это единственный процесс, поставляющий энергию. Гликолиз даёт большое кол-во энергии для обеспечения функций в анаэробных условиях. В 1ой стадии затрачивается 2 молекулы АТФ (гексокиназная и фосфофруктокиназная реакции). Во второй стадии образ 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная р-ии). Т.е. энергетич эффективность гликолиза сост 2 молеклы АТФ на 1 молекулу глюкозы. Биологич роль гликолиза: 1 Единств источник энергии в анаэробных условиях 2 Поставляет субстрат в ЦТК для полного расщепления глюкозы до воды и угл газа. 3 Явл источником субстратов для других биосинтезов (липидов, АК, глякозы). |
10. Значение ферментов гликолиза для эритроцитов. Гликолиз – единств процесс, продуцирующий АТФ в эритроцитах и поддерживающий их целостность и функции. Наследственный дефект пируваткиназы сопровождается гемолитической анемией. При этой патологии эритроциты имеют от 5% до 25% нормальной пируваткиназной активности, Следовательно, скорость глаколиза низкая. Промежуточный продукт гликолиза в эритроцитах – 2,3-дифосфоглицерат (2,3-ДФГ) – понижает сродство гемоглобина к кислороду. При недостаточности пируваткиназы сод-ние 2,3-ДФГ вдвое превышает норму, что обусловливает низкое сродство гемоглобина к кислороду. При недостаточности гексокиназы наблюдается понижение уровня 2,3-ДФГ и ненормально высокое сродство гемоглобина к кислороду. |
|
|
11. Киназные реакции гликолиза. Механизм гликолитической оксидоредукции. Регуляция гликолиза. К киназным реакциям относятся: фосфоглицераткиназная и пируваткиназная. Но кроме этих 2х сущ ещё две киназные реакции, но в них АТФ не образуется, а расходуется: гексокиназная и фосфофруктокиназная. Киназные реакции гликолиза являются фосфотрансферазными, то есть осуществляется перенос макроэргического фосфата с субстрата на АДФ в фосфоглицераткиназной и пируваткиназной реакциях и с АТФ на субстрат в гексокиназной и фосфофруктокиназной. Механизм гликолитической оксидоредукции. 2 гидрогиназные реакции гликолиза образуют сопряжённую пару и катализируют процесс, который называется гликолитической оксидоредукцией. 3ФГА → 1,3 ДФГК → 3 ФГК → 2 ФГК → → ФЕП → ПВК → ЛАКТАТ Лактат выполняет роль конечного ацептора протонов в ДЦ. РЕГУЛЯЦИЯ ГЛИКОЛИЗА Различают местную и общую регуляцию. Местная регуляция осуществляется путём изменения активности ферментов под действием различных метаболитов внутри клетки. Регуляция гликолиза в целом, сразу для всего организма, происходит под действием гормонов, которые, влияя через молекулы вторичных посредников, изменяют внутриклеточный метаболизм. Важное значение в стимуляции гликолиза принадлежит инсулину. Глюкагон и адреналин являются наиболее значимыми гормональными ингибиторами гликолиза. Инсулин стимулирует гликолиз через: активацию гексокиназной реакции; стимуляцию фосфофруктокиназы; стимуляцию пируваткиназы. на гликолиз влияют и другие гормоны. Например, соматотропин ингибирует ферменты гликолиза, а тиреоидные гормоны являются стимуляторами. Регуляция гликолиза осуществляется через несколько ключевых этапов. Реакции, катализируемые гексокиназой (1), фосфофруктокиназой (3) и пируваткиназой (10) отличаются существенным уменьшением свободной энергии и являются практически необратимыми, что позволяет им быть эффективными точками регуляции гликолиза.
|
12. Отличие реакций субстратного фосфорилирования от окислительного фосворилирования. Окислителное фосфорилирование: процесс образования АТФ, протекает в ДЦ митохондрий на внутренней мембране за счёт энергии трансмембранного протонного потенциала (восстановление НАД, НАДФ) Субстратное фосфорилирование 6 процесс образования АТФ, при котором активная фосфатная группа для фосфорилированной АТФ переносится непосредственно с субстрата. (цикл Кребса). |
13. Механизм окислительного декарбоксилирования ПВК. Строение полиферментного комплекса. Физиологическое значение. В аэробных условиях пировиноградная кислота подвергается окислительному декарбоксилированию с образованием ацетил-КоА. Это превращение катализируется надмолекулярным пируватдегидрогеназным комплексом, локализованным в матриксе митохондрий. В состав пируватдегидрогеназного комплекса входят три различных фермента: пируватдекарбоксилаза, дигидролипоатацетилтрансфераза и дегидрогеназа дигидролипоевой кислоты, их количественные соотношения в составе комплекса зависят от источника выделения, как правило это соотношение приближается к 30:1:10. Е1 катализирует декарбоксилирование ПВК с участием кофермента ТПФ. Образовавшийся продукт реакции при участии Е2 реагирует с окисленной липоевой кислотой. Липоевая кислота – низкомолекулярное азотсодержащее соединение явл коферментом Е2. Дисульфидная группа липоевой кислоты способна восстанавливаться и ацетилироваться. В реакции, катализируемой Е2, образ ацетиллипоевая кислота. Далее это соединение реагирует с коэнзимом А – при этом образуется восстановленная форма липоевой кислоты и ацетил-КоА. Далее начинает функционировать Е3, коферментом которого явл ФАД. Кофермент окисляет дигидролипоевую кислоту и сам при этом восстанавливается (ФАДН2). Восстановленый флавиновый кофермент реагирует с митохондриальным НАД+, восстанавливая его НАДНН+.
|
14. Аэробное расщепление углеводов в ЦТК.Реакции,ферменты,энергетический баланс.Эффект Пастера. За один оборот цикла Кребса происходит полное окисление одной молекулы ацетил – КоА . . Из четырёх пар атомов Н2 , три пары переносятся через НАД , а одна пара – через ФАД . На каждую пару атомов Н2 в системе биологического окисления образуется 3 ТФ ( 1 НАД * Н2 = 3 АТФ) .Следовательно , всего 9 АТФ . Одна пара атомов иона попадает в систему биологического окисления через ФАД – в результате образуется 2 АТФ . Кроме этого в ходе сукцинатилокиназной реакции образуется 1 ГТФ = 1 АТФ . Поэтому в ходе цикла Кребса образуется 12 АТФ . Функции цикла Кребса :1) Энергетическая ( цикл Кребса – конечный этап биологического окисления , в котором окисляется унифицированные соединения различного происхождения .2) Пластическая ( поскольку цикл « питается » субстратами различного происхождения , то он может быть источником углеродных скелетов для различных веществ ) 3) Регуляторная . Эффект Пастера-снижение скорости потребления глюкозы и прекращение накопления лактата в присутствии кислорода.ЭП-переход в присутствии кислорода от анаэробного гликолиза или брожения к дыханию.Механизм:конкуренция между системами дыхания и гликолиза(брожения) за АДФ,используемый для образования АТФ;уменьшение количества АДФ и соответствующее увеличение количества АТФ ведут к подавлению анаэробного гликолиза.
|
15.Неокислительные реакции ЦТК.Ферменты. 3я:Реакция катализируется НАД+–зависимым ферментом изоцитратдегидрогеназой. Фермент нуждается в присутствии ионов марганца (или Mg2+). Являясь по своей природе аллостерическим белком, изоцитратдегидрогеназа нуждается в специфическом активаторе – АДФ.4ая: Процесс катализируется α-кетоглутаратдегидрогеназой – ферментным комплексом, по стуктуре и механизму действия похожим на пируватдегидрогеназный комплекс.6ая: Фермент осуществляет прямой перенос водорода с субстрата (сукцината) на II комплекс дыхательной цепи митохондрий. Коферментом в этой реакции является ФАД.8ая: Реакция катализируется малатдегидрогеназой, коферментом которой служит НАД+.
|
|
|
16.Окислительные реакции ЦТК.Биологическое значение. Цикл трикарбоновых кислот – универсальный компонент биологического окисления , который базируется на принципе унификации. Унификация позволяет уравновешивать и оптимизировать соотношение основных субстратов , то есть если имеется избыток углеводов , то часть их перекачивается в липиды , если белка – то тоже в липиды или углеводы . Функции цикла Кребса :1) Энергетическая ( цикл Кребса – конечный этап биологического окисления , в котором окисляется унифицированные соединения различного происхождения .2) Пластическая ( поскольку цикл « питается » субстратами различного происхождения , то он может быть источником углеродных скелетов для различных веществ ) 3) Регуляторная .
|
17.ЦТК,ферменты,коферменты,витамины,регаляция. Первым регуляторным фактором является концентрация ЩУК , которая в основном образуется из ПВК . ПВК же образуется из углеводов ( глюкозы ) , поэтому при диабете или углеводном голодании наблюдается недостаток ПВК , а значит и ЩУК и сам цикл блокируется . АцетилКоА не является лимитирующим субстратом , так как в основном образуется при окислении жирных кислот . Но в то же время ЩУК – конкурентный ингибитор сукцинатдегидрогеназы , поэтому при избытке ЩУК цикл трикарбоновых кислот блокируется на 6 – ой стадии . Это торможение можно убрать глутамином , который переаминирует ЩУК в аспарагин . Второй регуляторный центр – концентрация НАД и НАД * Н2 . В живых системах концентрация НАД и Над * Н2 = const . Любые факторы , ведущие к увеличению НАД * Н2 ( алкогольная интоксикация ) и дефекту НАД блокируют цикл Кребса . Следовательно , увеличение концентрации НАД стимулирует цикл трикарбоновых кислот . Так как АТФ является косвенно конечным продуктом , то её избыток блокирует реакции , а значит АДФ стимулирует цикл ( АДФ рассматривается как аллостерический активатор изоцитратдегидрогеназы ) . Стимулятором Цикла является также и кислород , так как стимулирует работу АТФ . При повышении концентрации Са в клетке активируются дегидрогеназные реакции . Цикл Кребса активируется при сердечной недостаточности . Это объясняется тем , что миокард не может самостоятельно убрать избыток Са и эту роль берут на себя митохондрии , возрастает потребность в кислороде
|
18.Спиртовое брожение.Реакции.Сходство и отличие от гликолиза. В тканях различных организмов имеются ферменты , расщепляющие глюкозу до этанола . Этот процесс называется спиртовым брожением . По своему механизму спиртовое брожение очень близко к гликолизу . Расхождение начинается лишь после образования ПВК . При гликолизе ПВК под действием лактатдегидрогеназы превращается в лактат , а при спиртовом брожении этот конечный этап заменён двумя реакциями : пируватдекарбоксилазой и алкогольдегидрогеназой .
|
19.Метаболизм этанола.Механизм токсического действия на организм. Три пути метаболизма:а)алкогольдегидрогеназная реакция ( активность АДГ зависит от концентрации этанола в тканях ) + НАД ; АДГ СН3СН2ОН ------------- > СН3СОН -НАД*Н2 центральная фигура токсических эффектов этанола О НSКоА // СН3СОН СН3С --- цикл Кребса \ Н SКоА Н | |
СН3
– С = О + Н2N
– R
----
СН2 – О = N
–R
- Н2О Метаболические последствия этанольной интоксикации . Этанол монополизирует основной фонд НАД , переводя его в НАД*Н . Это приводит к блокированию первого комплекса дыхательной цепи и клетки , чтобы не погибнуть от гипоксии вынуждены переключиться на ФАД – зависимое окисление. Введение этанола приводит к сдвигам в нервной системе, которые обусловлены образованием продуктов конденсации СН3СОН с биогенными аминами. При приёме этанола повышается активность каталазы, которая провоцирует перекисные процессы , в результате развивается патология печени , сердца , нервной системы . Введение этанола создаёт экстремальную ситуацию , которая сопровождается выбросом адреналина , а значит и активацией липолиза и повышения содержания жирных кислот , которые поступают в печень . Жирные кислоты должны окисляться под действием НАД – зависимых дегидрогеназ , но эти реакции блокируются и получается , что печень просто не успевает их утилизировать и жирные кислоты накапливаются в печени . Второй причиной является образование ацетил – КоА из этанола , который тоже идётна синтез жирных кислот.
|
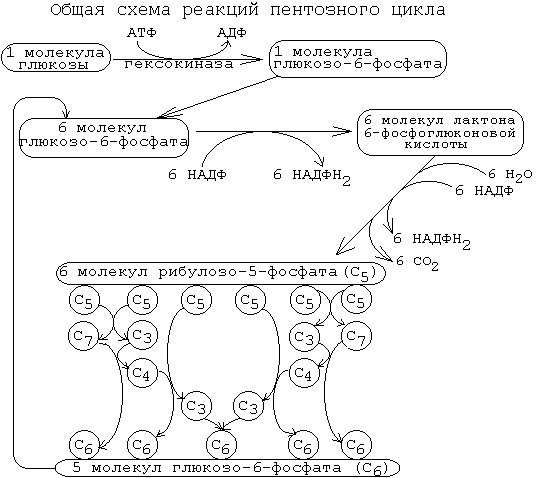
20)Роль тиамина в углеводном обмене II - НЕОКИСЛИТЕЛЬНЫЙ ЭТАП. Совокупность большого количества обратимых реакций. Каждая из них - это перенос 2-х или 3-х углеродного фрагмента с одного моносахарида на другой. Между моносахаридами происходит взаимный обмен частями своих молекул. При этом из пентозофосфатов, вступающих в реакцию, образуются моносахариды с разным числом углеродных атомов. Это триозы (например, фосфоглицериновый альдегид (ФГА)), тетрозы, гексозы, гептозы (их формулы знать необязательно, но нужно знать схему реакций. Реакции неокислительного этапа катализируются ферментами трансальдолазами и транскетолазами. В состав кофермента транскетолаз входит витамин В1 (тиамин). В результате 6 молекул рибозо-5-фосфата превращаются в 5 молекул глюкозо-6-фосфата.
|
|
|
21пентозный цикл. реакции, ферменты, локализ-ия в клетках и тканях ,физиологическое значение
Пентозофосфатный цикл активно функционирует в печени, жировой ткани, коре надпочечников, семенниках и в молочной железе в период лактации. В этих тканях активно идут процессы синтеза высших жирных кислот, аминокислот или стероидов, нуждающиеся в восстановительных эквивалентах в виде НАДФН+Н+.Цикл интенсивно работает также в эритроцитах, в которых НАДФН+Н+ используется для подавления перекисного окисления мембранных липидов. Мышечная ткань содержит очень малые количества глюкозо-6-фосфатдегидрогеназы и 6-фосфоглюконатдегидрогеназы, тем не менее, она также способна синтезировать необходимую клеткам рибозу.
|
22)субстратное,гормональное и энергетическое обюеспечение ГНГ субстраты ГНГ(ПВК,АК,глицерол,лактат) 1)глицеролглицерол-3-рдиоксиацетон-рфруктозо-1,6-дифосфатглюкоза 2)аланин+лактатпвкщукфосфоенол пвк3фгафруктозо-1,6-дифосфатглюкоза Гормональная регуляция. Повышение уровня глюкозы в крови приводит к выбросу инсулина α-клетками поджелудочной железы. Инсулин стимулирует поступление глюкозы в ткани и утилизацию её снижение концентрации глюкозы в крови. Гипогликемия провоцирует поступление в кровоток глюкагона и глюкокортикоидов, которые являются мощными активаторами ГНГ повышение концентрации глюкозы в крови (рис 6.11) ГНГ требует затрат энергии и протекает в условиях, при которых поддерживается достаточный уровень АТФ в клетке. повышение концентрации цАМФ в печени обуславливается действием глюкагона и адреналина; напротив, под действием инсулина она снижается. Суммарное уравнение глюконеогенеза из лактата: 2 лактат + 4 АТФ + 2 ГТФ -------> Глюкоза + 4 АДФ6 Н2О -------> 2 ГДФ + 6 Ф Из уравнения следует, что на синтез 1 молекулы глюкозы из 2 молекул лактата клетка затрачивает 6 макроэргических эквивалентов |
23)реакции ГНГ,ферменты,биологическое значение Глюконеогенез – механизм синтеза глюкозы.На образование 1 молекулы глюкозы затрачивается 2 молекулы ПВК.ГНГ протекает в печени и корковом веществе почек. В сущности, реакции ГНГпротекают в обратном направлении реакциям гликолиза – за одним важным исключением: требующие затрат энергии стадии гликолиза (гексокиназная, фосфофруктокиназная и пируваткиназная) не могут обратиться вспять.Для протекания глюконеогенеза требуется 4 фермента, не принимающих участия в окислительном расщеплении глюкозы - это пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатазаВ глюконеогенезе – 11 реакций, однако только 4 из них (пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, бифосфатаза и глюкозо-6-фосфатаза) считеются истинными фермнтами этого пути.Пируват является конечным продуктом гликолиза и исходной точкой глюконеогенеза. Биологическая роль глюконеогенеза: *переработка лактата *препятствует развитию лактат-ацидоза. *синтез глюкозы
|
24.особенности ГНГ в печени и почках,регуляция ГНГ Только в печени есть ещё один фермент, катализирующий реакцию превращения глюкозы в глюкозо-6-фосфат. Это изофермент гексокиназы - глюкокиназа. Гормональная регуляция. Повышение уровня глюкозы в крови приводит к выбросу инсулина α-клетками поджелудочной железы. Инсулин стимулирует поступление глюкозы в ткани и утилизацию её клетками организмаснижение концентрации глюкозы в крови. Гипогликемия провоцирует поступление в кровоток глюкагона и глюкокортикоидов, которые являются мощными активаторами глюконеогенеза. Следствием стимулированного глюконеогенеза становится повышение концентрации глюкозы в крови.Внутриклеточная регуляция. осуществляется путём аллостерического контроля ферментов, которые для этих двух путей различны.1)Активности гексокиназы и глюкозо-6-фосфатазы регулируются уровнем глюкозо-6-фосфата: гексокиназа им ингибируется, а фермент .2)Для пируваткиназы и пируваткарбоксилазы аллостерческим эффектором является ацетил-КоА, 3)Главным аллостерическим регулятором двух взаимосвязанных путей – гликолиза и глюконеогенеза – является фруктозо-2,6-дифосфат: увеличение его концентрации активирует ключевой фермент гликолиза – фосфофруктокиназу-1; снижение его концентрации активирует конкурирующую реакцию – образование фруктозо-6-фосфата, т.е. приводит к усилению глюконеогенеза |
25)цикл кюри и фелига между гликолизом, протекающим в мышцах при их интенсивной работе, и глюконеогенезом, осуществляемым печенью, существует тесная взаимосвязь (цикл Кори): образующая в мышцах молочная кислота поступает в общий кровоток, захватывается печенью и используется ею в качестве субстрата глюконеогенеза; синтезируемая при этом глюкоза отдаётся в кровототок и метаболизируется мышцами для получения энергии. Цикл Кори:
Цикл Фелига: Вместо лактата – аланин: в мышцах ПВК + NH2=аланин В печени:аланин – NH2= ПВК
|
|
|
26. Нормо-, гипо- и гипергикемия.Причины возникновения и клинические проявления.
Нормогликемия-нормальное содержание уровня глюкозы в крови. Гипогликемия-понижение концентрации глюкозы в крови. Гипергликемия- повышенное содержание концентрации глюкозы в крови.Гипергликемия может возникнуть при заболеваниях поджелудочной железы,в результате расстройства функции других эндокринных желез,участвующих в регуляции углеводного обмена.Поддержание постоянства уровня глюкозы в крови- это важнейшая функция печени, поэтому гипергликемия, обусловленная нарушением фунункции печени,выявляется при тяжелых ее поражениях.В клинике для изучения реактивности организма используют сахарную нагрузку. Гипогликемия связана с понижением тех эндокинных желез,повышение функций которых приводит к гипергликемии..Также может возникнуть при введении больным сахарным диабетом больших доз инсулина, при этом гипогликемия сопровождает почечную глюкозурию, возникающую вследствие снижения «почечного порога» для глюкозы. При нормальном уровне глюкозы в крови почки полностью её реабсорбируют и сахар в моче не определяется. Однако если гликемия превышает 9-10 ммоль/л (почечный порог), то появляется глюкозурия. При некоторых поражениях почек глюкоза может обнаруживаться в моче и при нормогликемии. В норме содержание глюкозы в крови натощак обычно ниже 6 ммоль/л, уровень в пределах 6-8 ммоль/л должен рассматриваться как пограничное состояние, а равный или превышающий 8 ммоль/л может служить диагнозом сахарного диабета.
|
27.Механизм срочной регуляции уровня глюкозы в крови. Роль ЦНС и гормонов(аденилатциклазный механизм).
Уровень глюкозы в крови является одним из гомеостатических параметров. Регуляция уровня глюкозы в крови – это сложный комплекс механизмов, обеспечивающий постоянство энергетического гомеостаза для наиболее жизненно важных органов (мозг, эритроциты). Глюкоза – главный и едва не единственный субстрат энергетического обмена. Существует два механизма регуляции: 1. Срочный (через ЦНС) 2. Постоянный (через гормональное влияние). Срочный механизм срабатывает практически всегда при действии на организм любых экстремальных факторов. Он осуществляется по классической модели (через зрительный анализатор воспринимается информация об опасности. Возбуждение из одного очага в коре распространяется по всем зонам коры. Затем возбуждение передаётся на гипоталамус, где находится центр симпатической нервной системы. По спинному мозгу импульсы поступают в симпатический ствол и по постганглионарным волокнам к коре надпочечников. При этом происходит выброс адреналина, который запускает аденилатциклазный механизм мобилизации гликогена). Срочный механизм поддерживает стабильную гликемию на протяжении 24 часов. В дальнейшем запас гликогена уменьшается и уже спустя 15 – 16 часов подключается постоянный механизм. |
28.Постоянный механизм регуляции уровня глюкозы в крови. Межорганный обмен субстратами. Основные гормоны,субстраты.
Уровень глюкозы в крови является одним из гомеостатических параметров. Регуляция уровня глюкозы в крови – это сложный комплекс механизмов, обеспечивающий постоянство энергетического гомеостаза для наиболее жизненно важных органов (мозг, эритроциты). Глюкоза – главный и едва не единственный субстрат энергетического обмена. Постоянный (через гормональное влияние): в основе которого лежит глюконеогенез. После истощения запасов гликогена, возбуждённая кора продолжает посылать импульсы в гипоталамус. Отсюда выделяются либерины, которые с током крови заносятся переднюю долю гипофиза, которая, в свою очередь, синтезирует в кровоток СТГ, АКТГ, ТТГ, которые в свою очередь стимулируют выброс трийодтиронина и тиреотропина. Эти гормоны стимулируют липолиз. Тиреотропные гормоны активируют протеолиз, в результате чего образуются свободные аминокислоты, которые как и продукты липолиза используются как субстраты глюконеогенеза и цикла трикарбоновых кислот. В ответ на повышение уровня глюкозы в крови, происходит выброс инсулина, однако вследствие того, что жирные кислоты и выделяемые гормоны выключают гликолиз в мышечной ткани, потребление глюкозы мышцами не происходит, вся глюкоза сохраняется для мозга и эритроцитов. В условиях длительного воздействия отрицательных факторов на организм (постоянный стресс) может возникнуть дефицит инсулина, что является одной из причин сахарного диабета.
|
29.Сахарный диабет 1 типа.Причины его возникновения(Механизм нарушений метаболизма). Диабет - это выделение глюкозы с мочой. диабет первого типа (диабет молодых людей, он является инсулиндефицитным). Диабет первого типа возникает в результате абсолютного или относительного дефицита инсулина. Абсолютный дефицит может быть вызван повреждением поджелудочной железы или разрушением в – клеток или островков Лангерганса, что возникает при опухолях, кистозах, травмах, хирургических вмешательствах. Относительный дефицит возникает при несоответствии между инсулиновой продукцией и его потреблением, то есть инсулин выделяется в недостаточном количестве. Причины: генетические дефекты молекулы инсулина, структуры белков – рецепторов, блокировки инсулина, относительного дефицита инсулина. Механизм: Срочный механизм осуществляется на первых этапах: происходит гиперполяризация мембраны, стимулируется работа Nа – К – АТФазы и создаётся потенциал, необходимый для транспорта субстратов для пластических и энергетических нужд. При дефиците инсулина все пути метаболизма глюкозы блокируются, так как возникает недостаточность глюкозы – 6 – фосфат. Дефицит инсулина снижает метаболизм глюкозы и аминокислот в тканях, это снимает инсулиновый тормоз с катаболических реакций: активизируется липолиз, протеолиз, гликогенолиз. Из – за дефицита глюкоза – 6 – фосфата в мышечной ткани нарушается производство АТФ и ткань находится в состоянии энергетического голода. Это состояние расценивается организмом как снижение уровня глюкозы в крови.
|
30.Биохимические изменения, возникающие в организме при недостаточности инсулина( гипеггликемия, глюкозурия, кетоз, гиперосмолярность).
Диабет второго типа: Эта форма связана с резистентностью инсулиновых рецепторов к инсулину. При этом типе Д происходит разрушение бета- клеток в рез-те аутоиммунных реакций. Биохимические изменения: повышенная концентрация в крови ЛПОНП, СЖК и главное кетоновых тел. Повышенное содержание ЖК в крови ведет к поглощению их печенью, где из них синтезируются ТГ ( в адипоцитах) , который далее в составе ЛПОНП секретируются в кровь. Другая часть ЖК вступает на путь бета- окисления в митохондриях печени, образуется СН3-СО-SКоА, из которого даоее синтезируются кетоновые тела. При СД возможны 3 коматозных состояния: кетоацидотическая кома с абсолютной ИНС недостаточностью; гиперосмолярная кома с умеренной недостаточностьюИНС;лактатацидотическая кома с выраженной гипоксией, сепсисом, СС шоком. При инсулинотерапии может быть гипогликемическая кома, связанная с передозировкой ИНС. Первые три состоянии возможны не только при СД, но и при действии многих др.факторов. Три основные формы коматозных состояний никогда не встречаются в отдельности. Обычно проявляется какая-то из низ, обычно гиперосмолярный синдром. В период комы С-пептид и иммунореактивный ИНС в крови не определяются, гипергликемия отмечается всегда.Ацидоз при диабетической коме- это накопление органических кислот: кетоновых тел, лактата, пирувата, конц-я кетоновых тел в 200 раз превышает норму. Она повышается как следствие повышенного синтеза кетоновых.тел в печени, но и является результатом олигурии и анурии, которые часто бывают при коме. рН крови снижается.Глюкозурия - появление в моче сахара. |
|
|
31.Механизм кетонурии и кетонемии при голодании, и при сахарном диабете.
Кетонемия-повышение содержания кетоновых тел в крови до 20 ммоль/л.Кетонемия сопровождается резким увеличением кетоновых тел в моче- кетонурия.Явление кетонурии и кетонемии объясняется следующим образом. И диабет, и голодание сопровождаются резким сокращением запасов гликогена в печени. Многие ткани и органы, в частности мышечная ткань, находятся в состоянии энергетического голода (при недостатке инсулина глюкоза не может с достаточной скоростью поступать в клетку). В этой ситуации благодаря возбуждению метаболических центров в ЦНС импульсами с хе- морецепторов клеток, испытывающих энергетический голод, резко усиливаются липолиз и мобилизация большого количества жирных кислот из жировых депо в печень. В печени происходит интенсивное образование кетоновых тел. Образующиеся в необычно большом количестве кетоновые тела с током крови транспортируются из печени к периферическим тканям. Периферические ткани при диабете и голодании сохраняют способность использовать кетоновые тела в качестве энергетического материала, однако ввиду необычно высокой концентрации кетоновых тел в притекающей крови мышцы и другие органы не справляются с их окислением и как следствие возникает кетонемия. |
32.Диагностика сахарного диабета.Техника построения «сахарной» кривой. Клинические проявления СД. При нормальном уровне глюкозы в крови почки полностью её реабсорбируют и сахар в моче не определяется.,если гликемия превышает 9-10 ммоль/л, то появляется глюкозурия. При некоторых поражениях почек глюкоза может обнаруживаться в моче и при нормогликемии. В норме содержание глюкозы в крови натощак обычно ниже 6 ммоль/л, уровень в пределах 6-8 ммоль/л должен рассматриваться как пограничное состояние, а равный или превышающий 8 ммоль/л может служить диагнозом сахарного диабета. Проверка способности организма регулировать содержание глюкозы в крови используется для диагностики сахарного диабета при постановке перорального глюкозо-толерантного теста: Первая проба крови берётся натощак после ночного голодания. Затём больному в течение 5 мин. дают выпить раствор глюкозы (75г глюкозы, растворённой в 300 мл воды). После этого каждые 30 мин. на протяжении 2-х часов определяют содержание глюкозы в крови. Диагностика СД: Концентрация ГЛ больше 7. 2 ммоль/л указывает на СД. И нет необходимости проводить тест толерантности к глюкозе. Наличие гликозилированного Нb. При СД его концентрация увеличивается в 2-3 раза. Инс и С-пептид секретируются бета- клетками в эквимолярных количествах. В печени задерживается около 60% ИНс, поступающего с кровью воротной вены из поджелуд. железы. Поэтому отношение С-петид/ Инс в воротной вене и периф. кровообращении при н.ус. равно примерно—3/1. С-пептид удаляется обычно через почки его суточная секреция-45 мкг и пропорц. суточной секреции ИНС. Альбуминурия.- ранний признак СД. В норме с мочой-выводится в среднем 8мг А. При выделении 30-300мг, микроальбуминурия. |
33.Гликогеновын болезни
1.Гликогенозы 2.Агликогенозы Гликогенозы-болезни накопления 1-й тип: б-знь Гирке-недостаточность глюкозо-6 фосфатазы.наблюдается гепатомегалия.нарушение ф-ии почек и печени 2-й тип-б-знь Помпе-отсутствие лизосомальной глюкозидазы.при этом-генерализованное поражение органов. 3-й тип:б-знь Форбса-дефект деветвящегося фермента.Накопление полисахарида 4-й тип:б-знь Андерсена-дефект фермента ветвления-лет.исход в 1-й год жизни от недостаточности 5-й тип:б-знь Мак-Ардла-недостаточность мышечной фосфорилазы 6-й тип:б-знь Херса-недостаточность фофсфорилазы в печени-развивается гепатомегалия. При агликогенозах в результате нарушения синтеза гликогена страдают энергетические ресурсы клетки. |
34. Глюкуроновая кислота используется для синтеза ГАГ.УДФ-глюкуроновая к-та,связывая продукты метаболизма и чужеродные вещ-ва легко экскретируется в желчью и мочой. Синтез: глюкозо-6 фосфат+фосфоглюкомутаза= глюкозо-1-фосфат+УТФ+УДФглюкозипиро фосфорилаза=УДФ-глюкоза окисляется дегидрогеназой в УДФ-глюкуроновую к-ту,теряет УДФ=глюкуроновая к-та
|
35.
Существует особый вид соединений –это белково-углеводные комплексы. Различают: 1)протеогликаны (свыше 95 %углеводов) 2)гликопротеины (до 10% углвеводов) ПГ-белковые комплексы,в кот. С молекулами белка ковалентно связаны гликозамингликаны. Ф-ии ПГ: структурный компонент внеклет.матрикса 2)связывает поликатионы и катионы 3) обеспечивает тургор тканей 4) влияет на клеточную миграцию 5)регулирует фильтрацию в клубочках почек 6)входит в состав везикул Ф-ии гликопротеинов: 1) структурный компонент мембраны Кл. 2) защитные св-ва (муцин) 3) иммунная защита (интерферон) 4) гормоны-гликопротеины |
|
|
36. Гликозамингликаны-полисахариды.Их цепи подвижны и не могут образовывать глобулы.Из-за гидрофильности занимают большие объёмы,образуя гели и создавая тургор тканей. Содержит сульфатированные сахара,имеет тенденцию состоять из разных дисахаридных единиц,синтезируются внутриклеточно и покидают клетку путём экзоцитоза; сахара ковалентно связаны с белками в форме ПГ ГАГ классифицируют: 1)гиалуронаты 2)хондроитин- и дерматансульфаты 3)кератинсульфаты 4) гепарин и гепарансульфаты |
37. Мукополисахаридозы - группа наследственных болезней соединит. ткани, обусловленных нарушением обмена гликозаминогликанов в результате генетически обусловленной неполноценности ферментов, участвующих в их расщеплении. Наследуется по аутосомно-рецессивному типу. При этом поражается система лизосомных ферментов, участвующих в катаболизме гликозаминогликанов. Вследствие ферментативной недостаточности последние накапливаются в большом количестве в органах и тканях, поэтому мукополисахаридозы относят к болезням накопления. В результате нарушается функциональное состояние различных органов и систем, а поскольку гликозаминогликаны входят в состав соединительной ткани, то одним из ведущих проявлений мукополисахаридоза является системное поражение скелета, задержка физического развития. По современной классификации, в зависимости от характера ферментативного дефекта выделяют восемь осн. типов мукополисахаридозов. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|









 лицериновый
Диоксиацетонфосфат
лицериновый
Диоксиацетонфосфат
 АТФ
АТФ

 Галактоза
галактоза-1-Ф
УДФ-глюкоза
Галактоза
галактоза-1-Ф
УДФ-глюкоза

 Гл-1-Ф
УДФ-галактоза
Гл-1-Ф
УДФ-галактоза


 УДФ-глюкоза
УДФ-глюкоза