

Биофизика 02
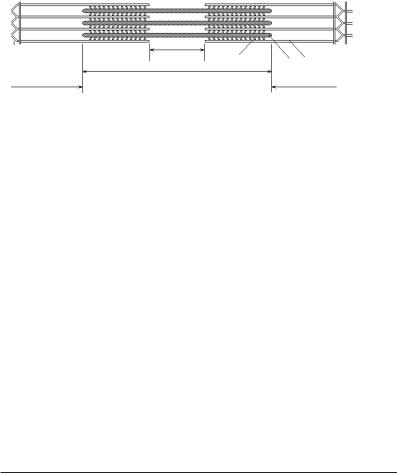
.pdfфрагменти: легкий мероміозин (ЛММ)
іважкий мероміозин (ВММ), між ними є шарнір. ВММ складається з двох
субфрагментів: S1 і S2. ЛММ і фрагмент S2 ВММ вкладені в пучок ниток, а субфрагмент S1 виступає над поверхнею. Цей виступаючий кінець (міозинова головка) здатний зв’язуватися з активним центром на актиновій нитці
ізмінювати кут нахилу до пучка міозинових ниток.
Об’єднання окремих молекул міозину в пучок (рис. 7.4) відбувається, швидше за все, за рахунок електро-
статичних взаємодій між ЛММ. Центральна частина нитки (біля 300 нм) не має головок. Весь комплекс міозинових молекул простирається на 1,5 мкм. Це одна з найбільших молекулярних структур, відомих у природі.
При розгляданні в поляризаційному мікроскопі подовжнього зрізу поперечноFсмугастого м’яза видно світлі і темні ділянки. Темні ділянки (диски) є анізотропними: у поляризованому світлі вони виглядають прозорими в подовжньому напрямку і непрозорими у поперечному, вони позначаються літерою А. Світлі ділянки є ізотропними і позначаються літерою I (рис. 7.5). Диск I містить у собі тільки тонкі нитки, диск А — і товсті, і тонкі. У середині диска А видна світла смуга, називана НFзоною. Вона не має тонких ниток. Диск I розділений тонкою смугою Z. Ця смуга являє собою мембрану, що
містить структурні елементи, які скріплюють між собою кінці тонких ниток. Ділянка між двома ZFлініями називається саркомером.
Кожна товста нитка оточена шістьма тонкими, а кожна тонка — трьома товстими. Таким чином, у поперечному зрізі м’язове волокно має правильну гексагональну структуру (рис. 7.6).
141
§ 37. Скорочення м’яза
При скороченні м’яза довжина актинових і міозинових філаментів не змінюється. Відбувається лише їхній зсув щодо один одного: тонкі нитки всовуються в проміжки між товстими. При цьому довжина диска А залишається незмінною, а диск I укорочується, смуга Н майже зникає. При цьому довжина саркомера змінюється приблизно від 2,5 до 1,7 мкм.
Як уже вказувалося, міозинова нитка має на собі безліч головок, якими вона може зв’язуватися з актином. Актинова ж нитка, у свою чергу, має ділянки (активні центри), до яких можуть прикріплюватися головки міозину. У спочиваючій м’язовій клітині ці центри зв’язування прикриті молекулами тропоміозину, що перешкоджає утворенню зв’язку між тонкими і товстими нитками (рис. 7.7).
Z[лінія
I[диск
Н[зона |
3 |
1 |
2 |
|
|
|
А[диск
I[диск
Рис. 7.7. Будова поперечно[смугастого м’яза (подовжній зріз). 1 — товста (міозинова) нитка; 2 — тонка (актинова) нитка; 3 — поперечні містки, що забезпечують зв’язок між актиновими і міозиновими нитками.
Для того, щоб актин і міозин могли взаємодіяти, необхідна присутність іонів кальцію. У спокої вони знаходяться в так званому саркоплазматичному ретикулумі. Ця органела являє собою обмежені мембранами порожнини, що містять кальцієвий насос, який за рахунок енергії АТФ транспортує іони кальцію всередину саркоплазматичного ретикулума. Його внутрішня поверхня містить білки, здатні зв’язувати Са2+, що декілька зменшує різницю концентрацій цих іонів між цитоплазмою і порожниною саркоплазматичного ретикулума. Потенціал дії, що поширюється по клітинній мембрані, активує близько розташовану до поверхні клітини мембрану саркоплазматичного ретикулума, що викликає вихід Са2+ у цитоплазму.
Молекула тропоніну має високу спорідненість до кальцію. Під його впливом вона змінює положення тропоміозинової нитки на актиновій таким чином, що відкривається активний центр, раніше прикритий тропоміозином. До активного центру, що відкрився, тепер може приєднатися поперечний місток. Це приводить до взаємодії актину з міозином. Після утворення такого зв’язку міозинова головка, раніше розташована
142

майже під прямим кутом до |
|
4 |
2 |
ниток, нахиляється і протаскує |
а) |
1 |
3 |
актинову нитку щодо міозино- |
|
||
|
|
5 |
|
вої на 10 нм (рис. 7.8). |
|
|
|
|
|
|
|
АктинFміозиновий комп- |
б) |
|
|
лекс, що утворився, переш- |
|
|
|
коджає подальшому ковзанню |
|
|
|
ниток одна щодо одної, тому |
в) |
|
|
необхідно його роз’єднання. |
|
|
|
Це можливо лише за раху- |
г) |
|
|
нок енергії АТФ. Міозин має |
|
|
|
АТФазну активність, тобто |
|
|
|
здатний викликати гідроліз |
Рис. 7.8. Скорочення саркомера: 1 — міозинова |
||
АТФ. Енергія, що виділяється |
|||
при цьому, розриває зв’язок |
нитка; 2 — актинова нитка; 3 — міозинова голо- |
||
між актином і міозином. Після |
|
вка; 4 — активний центр; 5 — Z[лінія. |
|
а) взаємодія між тонкими і товстими нитками |
|||
цього міозинова головка здатна відсутня; б) у присутність Са2+ міозинова голо-
взаємодіяти з новою ділянкою |
вка зв’язується з активним центром на акти- |
|
молекули актину. |
новій нитці; в) поперечні містки нахиляються |
|
і протягують тонку нитку щодо товстої, у ре- |
||
При розслабленні м’яза ак- |
||
зультаті чого довжина саркомера зменшується; |
тивізується робота кальцієвого г) зв’язки між нитками розриваються за рахунок
насоса, що знижує концент- |
енергії АТФ, міозинові головки готові взаємодія- |
рацію Са2+ у цитоплазмі, отже |
ти з новими активними центрами. |
зв’язки між тонкими і товстими |
На рисунку для простоти показані тільки два |
містки. У реальному м’язі їх набагато більше. |
|
нитками вже не можуть утво- |
Їхня робота синхронізована таким чином, що |
рюватися. У цих умовах при |
зв’язування, нахил і розриви всіх містків однієї |
розтягу м’яза нитки безпере- |
нитки відбувається одночасно. |
шкодно сковзають одна щодо |
|
одної. Однак така розтяжність |
|
можлива тільки в присутності АТФ.
Якщо в клітині відсутня АТФ, то актинFміозиновий комплекс не може розірватися. Нитки залишаються жорстко зчепленими між собою. Це явище спостерігається при трупному задубінні.
Розрізняють два режими скорочення м’яза: ізотонічний, при якому змінюється довжина волокна, а напруга залишається незмінною, і ізометричний, при якому кінці м’яза нерухомо закріплені, унаслідок чого змінюється не довжина, а напруга.
§ 38. Потужність і швидкість скорочення м’яза
Важливими характеристиками роботи м’яза є сила і швидкість скорочення. Рівняння, що виражають ці характеристики, були емпірично отримані А. Хіллом і згодом підтверджені теоретично кінетичною тео-
143

рією м’язового скорочення.
Рівняння Хілла, що зв’язує між собою силу і швидкість скорочення м’яза, має наступний вигляд:
(P + a)(V + b) = (P0 + a)b = a(Vmax + b),
де V — швидкість укорочення м’яза; Р — м’язова сила або прикладене до неї навантаження (зовнішня сила, що діє на м’яз); Vmax — максимальна швидкість
скорочення м’яза (V = Vmax при P = 0); P0 — сила, що розвивається м’язом в ізомет-
ричному режимі скорочення, тобто при такому навантаженні, при якому не відбувається зміна довжини м’яза (P =P0 при V =0); a і b — константи. Приведене рівняння має вигляд гіперболи (рис. 7.9).
При скороченні за час t м’яз робить роботу А:
У процесі скорочення м’яза виділяється деяка кількість теплоти Q. Ця величина називається теплопродукцією. Як показав Хілл, теплопродукція залежить лише від зміни довжини х м’яза і не залежить від навантаження Р :
Q = ах. |
(7.1) |
Потужність N, що розвивається м’язом, визначається швидкістю чинення роботи і виділення теплоти:
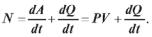 (7.2) Підставивши вираз (7.1) у (7.2), одержимо наступне:
(7.2) Підставивши вираз (7.1) у (7.2), одержимо наступне:
або, як випливає з рівняння Хілла,
N = (Р + а)V = b(P0 –P),
тобто залежність потужності N від навантаження P є лінійною (рис. 7.10).
Коефіцієнт корисної дії м’яза η записується у вигляді:
η зберігає постійне значення (біля 40%) у діапазоні значень сили від 0,2Р0 до 0,8Р0.
144
Константи а і b мають постійні |
|
||
значення для даного м’яза. Кон- |
|
||
станта а має розмірність сили, а |
|
||
b — швидкості. Константа b у знач- |
|
||
ній мірі залежить від температури, |
|
||
наприклад, при нагріванні на 10 °С |
|
||
поблизу 0 °С b подвоюється. Кон- |
|
||
станта а знаходиться в діапазоні |
|
||
значень від 0,25Р0 до 0,4Р0. За цими |
|
||
даними легко оцінити максимальну |
|
||
швидкість скорочення для даного |
|
||
м’яза. Так як |
|
||
|
|
|
Рис. 7.10. Залежність сумарної потуж- |
|
|
|
ності кравцового м’яза жаби від наван- |
то V |
max |
перевищує константу b у 2,5 |
таження при тетанічному ізотонічному |
|
|
скороченні. |
|
– 4 рази.
Приклади розв’язання задач
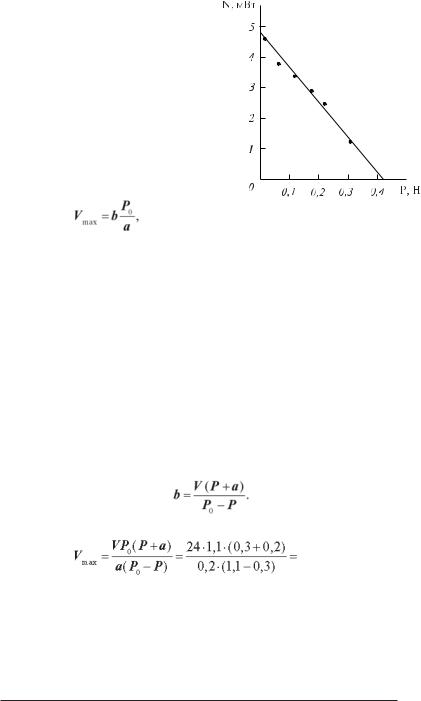
Задача 7.1. Для деякого м’яза при навантаженні 0,3 Н швидкість скорочення складає 24 мм/с. Навантаження в ізометричному режимі скорочення складає 1,1 Н, стала а дорівнює 0,2 Н. Обчислити максимальну швидкість скорочення.
Розв’язання. Запишемо рівняння Хілла
(P + a)(V + b) = (P0 + a)b = a(Vmax + b). З останньої рівності знайдемо Vmax:
Vmax = bP0 /a,
аз першої — константу b:
Тоді
72 мм/с.
Задача 7.2. М’яз, скорочуючись із швидкістю 6 мм/с, розвиває потужність 2,7 мВт. Навантаження в ізометричному режимі скорочення для цього м’яза складає 0,8 Н, константа b дорівнює 23 мм/с. Обчислити роботу, зроблену м’язом за одну секунду.
Розв’язання. Робота м’яза дорівнює
145

A = PVt.
Навантаження Р знайдемо з формули для потужності:
N = b(P0 – P),
звідки
Тоді
= 4,1·10–3 Дж.
Питання тестового контролю
7.1.До складу тонких ниток м’язового волокна входять: а) актин і тропонін; б) актин, міозин і тропоміозин; в) міозин;
г) актин, тропоміозин і тропонін; д) актин і міозин.
7.2.До складу товстих ниток м’язового волокна входять: а) актин і тропонін; б) актин, міозин і тропоміозин; в) міозин;
г) актин, тропоміозин і тропонін; д) актин і міозин.
7.3.При ізотонічному скороченні:
а) довжина волокна змінюється, напруга постійна; б) напруга змінюється, довжина волокна постійна; в) довжина і напруга незмінні; г) довжина і напруга змінюються.
7.4. При ізометричному скороченні:
а) довжина волокна змінюється, напруга постійна; б) напруга змінюється, довжина волокна постійна; в) довжина і напруга незмінні; г) довжина і напруга змінюються.
7.5. Рівняння Хілла має наступний вигляд: а) (P + a)(V + b) = (P0 + a)b = a(Vmax + b);
б)
в) (P + a)(V – b) = (P0 + a)b = a(Vmax – b);
146

г)
д) (P – a)(V + b) = (P0 + a)/b = a(Vmax – b).
7.6. Потужність N, що розвивається м’язом, дорівнює:
а) 
б) 
в) N = (Р – а)V = b(P0 + P);
г) 
д) N = (Р + а)V = b(P0 – P).
7.7. Коефіцієнт корисної дії м’яза η записується у вигляді:
а)
б) 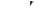
в) 
г) 
д)
147
Розділ 8. БІОФІЗИКА ЗОРУ
§ 39. Оптична система ока людини
Око має не зовсім правильну сферичну форму. Його вертикальний, горизонтальний і передньозадній розміри дорівнюють, відповідно, 23,4 мм, 23,6 мм і 24,3 мм.
Світло потрапляє в око через рогівку. Показник заломлювання її речовини 1,38, радіус кривизни — 7–8 мм. Ця частина ока має найбільші заломлюючі властивості. За рогівкою знаходиться кришталик, що являє собою двоопуклу лінзу з радіусом кривизни передньої поверхні 10 мм, задньої 6 мм. Показник заломлювання речовини кришталика складає 1,4. Простір між рогівкою і кришталиком заповнений водянистою вологою, а внутрішня частина ока — склоподібним тілом — прозорою гелеподібною масою, близькою за своїми оптичними властивостями, так само як і водяниста волога, до води. Внутрішній шар ока, що містить зорові елементи (див. нижче), називається сітківкою. До сітківки підходить зоровий нерв, що відводить нервові імпульси в зоровий центр головного мозку. У області прилягання зорового нерва до сітківки знаходиться сліпа пляма, нечутлива до світла, а всередині сітківки — сама чутлива до світла область — жовта пляма.
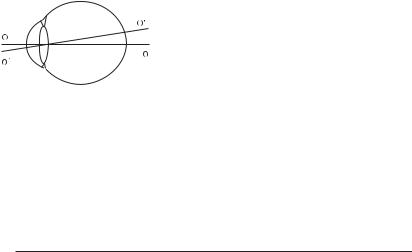
Через геометричний центр рогівки і кришталика проходить головна оптична вісь ОО, а через центр кришталика і жовту пляму — зорова вісь О’О’
(рис. 8.1). Зорова вісь збігається з на-
прямком найкращої світлочутливості.
Кут між головною оптичною і зоровою
осями складає приблизно 5°.
Око має чотири заломлюючі поверхні,
Рис. 8.1 (пояснення в тексті). |
утворені рогівкою, водянистою вологою і |
|
|
|
кришталиком. Зовні така оптична систе- |
ма обмежена повітрям, зсередини — склоподібним тілом. Для простоти всю оптичну систему ока представляють лінзою, що з боку простору предметів оточена повітрям, а з боку простору зображень — рідиною з показником заломлювання 1,38. Така спрощена система називається приведеним редукованим оком (рис. 8.2).
Показники заломлювання повітря і внутрішнього середовища ока нерівні. Наслідком цього є:
148
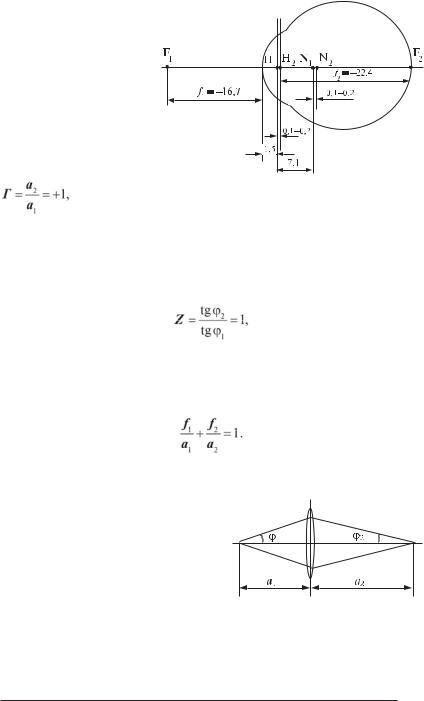
1.Оптичні відстані f1 і f2 нерівні (рис. 8.2).
2.Головні площини оптичної системи ока, які перпендикулярніголовній оптичній осі і проходять
через точки Н1 і Н2 (рис. 8.2), не збігаються. Н1 і Н2 — це точки, для яких лінійне збільшення
Рис. 8.2. Приведене редуковане око (пояснення в тексті, |
відстані — в мм). |
де а1 — відстань від предмета до лінзи; а2 — відстань від лінзи до зображення (рис. 8.3).
3. З головними площинами не збігаються площини, які перпендикулярні головній оптичній осі і проходять через точки N1 і N2 (рис. 8.2). Для цих точок кутове збільшення
де ϕ1 — кут розкриття пучка променів точки предмета; ϕ2 — кут розкриття пучка променів для сполученої точки зображення (рис. 8.3).
З геометричної оптики відомо, що
Тому для одержання чіткого зображення поFрізному віддалених предметів повинні змінюватися або відстань а2, або — фокусна відстань f2. В оці людини реалізується останній
спосіб, що є можливим завдяки зміні кривизни кришталика. Можливість фокусування на сітківці зображень поFрізному віддалених предметів називається акомодацією. Чим ближче
розташований предмет, тим більша кривизна кришталика і його оптична сила.
У здорової людини акомодація не викликає напруги при розгляданні предметів, що знаходяться на від-
стані більш, ніж 25 см. Ця найменша відстань називається відстанню
149
найкращого бачення.
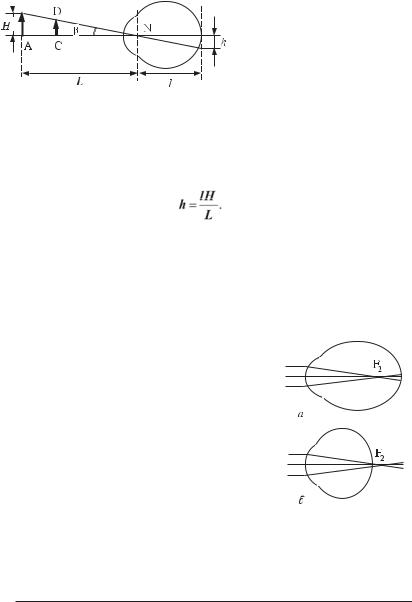
Чим більше предмет віддалений від ока, тим менше його зображення на сітківці. Проведемо промені через крайні точки предмета АВ, що має розмір H, і збіжну вузлову точку N (рис. 8.4). Перетинання їх із сітківкою дасть дійсне, зворотне, зменшене зображення предмета розміром h. Якщо кут зору 
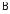
 β між променями малий, то
β між променями малий, то
отримаємо
Рис. 8.4. Побудова зображення предмета.
H = Lβ,
де L — відстань від предмета до ока. Тоді
h = lβ
де l — відстань між збіжною вузловою точкою N і сітківкою.
Розмір предмета H складає
З останнього виразу видно, що знання відстані до предмета дозволяє визначити розмір останнього, і, навпаки, знання розміру предмета дозволяє визначити відстань до нього.
Найменший кут зору β, при якому людина ще здатна бачити окремо дві різні точки предмета, складає приблизно одну хвилину. При розташуванні предмета на відстані 25 см розділювальна здатність людського
ока складає 70 мкм, а розмір зображення цих |
|
точок на сітківці буде складати 5 мкм, що, у свою |
|
чергу, дорівнює середній відстані між колбочка- |
|
ми (див. нижче). |
|
В нормі задній фокус оптичної системи ока |
|
повинен збігатися із сітківкою. При короткозо- |
|
рості внаслідок збільшення передньозаднього |
|
розміру очного яблука фокус розташований |
|
перед сітківкою, що викликає розмите зоб- |
|
раження далеко розташованих предметів |
|
(рис. 8.5,а). Для короткозорого ока відстань най- |
|
кращого бачення менша 25 см. Цей недолік зору |
|
коригується розсіювальними лінзами. |
Рис. 8.5. а) хід променів у |
При далекозорості, навпаки, очне яблуко |
короткозорому оці; б) хід |
зменшене, і промені фокусуються за сітківкою при |
променів у далекозорому |
розгляданні близько розташованих предметів (рис. |
оці. |
|
150