

Биофизика 02
.pdfБуло замічено, що амплітуда потенціалу дії не залежить від сили подразника, а сам потенціал виникає лише в тому випадку, якщо деполяризація перевищила деякий граничний рівень, обумовлений тільки властивостями даної клітини. Це явище одержало назву закону “усе або нічого”. Однак, якщо деполяризація складає 50–75% від рівня граничної, то в клітині може виникнути локальна відповідь, амплітуда якої значно нижча амплітуди потенціалу дії (рис. 6.4). Чим вищий рівень підпорогової деполяризації, тобто чим ближчий він до порогової, тим вища амплітуда локальної відповіді. 
Невиникнення потенціалу дії при підпороговому рівні
деполяризації пояснюють, |
Рис. 6.4. Локальна відповідь. А, Б, В — зміна |
|
трансмембранного потенціалу при дії підпорого- |
||
тим, що при цьому недостат- |
||
вого збудження; Б, В — підпорогова деполяризація |
||
ньо збільшується натрієва |
стимулювала розвиток локальної відповіді (пасивні |
|
проникність, щоб викликати |
зміни потенціалу позначені пунктирною лінією); |
|
регенеративну деполяри- |
Г — критичний рівень деполяризації переростає в |
|
потенціал дії. |
||
зацію. Виникаючий рівень |
||
|
||
деполяризації не викликає |
|
відкриття нових натрієвих каналів, тому натрієва провідність швидко зменшується, і в клітині відновлюється потенціал спокою.
Слід зазначити, що амплітуда потенціалу дії і граничний рівень деполяризації не є строго постійними величинами для даної клітини. Тривала деполяризація приводить до збільшення інактивації натрієвих каналів і активації калієвих, наслідком цього є зменшення амплітуди потенціалу дії і збільшення граничного рівня деполяризації. Тривала гіперполяризація викликає зворотні ефекти: збільшення амплітуди потенціалу дії і зменшення граничного рівня деполяризації.
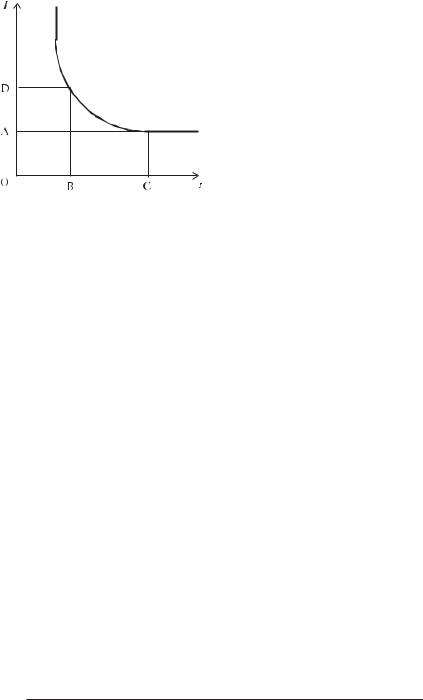
Виникнення потенціалу дії залежить не тільки від сили струму, але і від часу його дії. Чим більша сила струму, тим менше часу потрібно подавати його на клітину, щоб виник потенціал дії. Дія великих струмів протягом короткого проміжку часу не викликає потрібного рівня деполяризації, так само як і дія протягом тривалого часу токів малої сили. Найменший струм, що здатний викликати потенціал дії називається реобазою (відрізок ОА на рис. 6.5), час збудження, що відповідає йому
— корисним часом (відрізок ОС на рис. 6.5). У біології і медицині часто використовується термін “хронаксія” (відрізок OВ на рис. 6.5), що означає час, протягом якого повинен діяти струм у дві реобази (відрізок OD на рис. 6.5). Для кожної клітини існує своє значення реобази і хронаксії, тому вимір цих характеристик має діагностичне значення.
131
|
§ 34. Поширення збудження по |
||
|
нервовому волокну |
||
|
Розрізняють м’якотні (мієліни- |
||
|
зовані) і безм’якотні (немієлінизо- |
||
|
вані) нервові волокна. Мієлінизо- |
||
|
ване нервове волокно складається |
||
|
з осьового циліндра, який пок- |
||
|
ритий цитоплазматичною мемб- |
||
|
раною і містить аксоплазму (рис. |
||
|
6.6). Навколо нього багаторазово |
||
|
обертаються швановські клітини |
||
|
(у периферичній нервовій системі) |
||
Рис. 6.5. Крива сили — тривалості: |
або олігодендроглії (у центральній |
||
I — сила струму; t — час. |
нервовій системі), шари яких зли- |
||
|
ваються й утворюють мієлінову |
||
оболонку нервового волокна. Через рівні проміжки (від 0,2 до 2 мм), |
|||
характерні для даної клітини, ця оболонка переривається, і мембрана |
|||
осьового циліндра залишається відкритою. Такі ділянки волокна на- |
|||
зиваються перехватами Ранв’є. Їхня довжина становить приблизно 1 |
|||
мкм. Мієлінова оболонка, що складається з мембранних ліпідів і білків, |
|||
є надійним ізолятором нервової клітини, завдяки їй збудження може |
|||
виникнути тільки на оголеній ділянці мембрани аксона. Безм’якотні |
|||
нервові волокна не мають такої щільної жирової оболонки. Швановська |
|||
клітина оточує їх тільки один раз. |
|
||
Збудження якоїсь ділянки безм’якотного нервового волокна приво- |
|||
дить до локальної деполяризації мембрани, тобто в цій області зовнішнє |
|||
середовище стає зарядженим негативно, а внутрішнє — позитивно. У |
|||
той же час інша (незбуджена) частина мембрани зберігає свою звичай- |
|||
ну різницю потенціалів: зовнішнє середовище заряджене позитивно, |
|||
а внутрішнє — негативно. Між збудженою і незбудженою областями |
|||
виникають місцеві струми (рис. 6.7). Це приводить до деполяризації |
|||
сусідньої ділянки, що, у свою чергу, деполяризує наступний. Приєднав- |
|||
ши електроди осцилографа |
1 |
2 |
|
до двох ділянок клітинної |
|||
|
|
||
мембрани, можна спостері- |
|
|
|
гати поширення потенціалу |
3 |
|
|
дії. Такий спосіб проведення |
|
|
|
збудження є безперервним. |
4 |
5 |
|
У м’якотних нервових |
Рис. 6.6. Мієлінизоване нервове волокно: |
||
волокнах неможливе безпе- |
1 — швановська клітина; 2 — ядро швановської |
||
рервне проведення нерво- |
клітини; 3 — аксоплазма; перехват Ранв’є; |
||
вого імпульсу. Збудження |
5 — мієлін. |
|
|
|
|
||
132

(деполяризація) може виникати не по всій довжині мембрани, а тільки в перехватах Ранв’є. Деполяризація однієї такої ділянки А викликає деполяризацію сусідньої ділянки В (рис. 6.8). Далі збудження здатне перейти тільки до ділянки В, тому що ділянка А протягом деякого часу залишається
нечутливою до збудження (рефрактерною). З цієї причини імпульс поширюється по нервовому волокну тільки в одному напрямку. Виникаючий потенціал дії в кілька разів перевищує поріг, необхідний для в и н и к н е н н я з б у д ж е н н я в наступному перехваті Ранв’є, що, таким чином, щоразу підсилює сигнал, який слабшає в результаті опори міжклітинної рідини й аксоплазми.
Такий стрибкоподібний (сальтаторний) спосіб проведення збудження подібний дії ретранслюючого генератора. Стрибкоподібне проведення збудження є більш вигідним, тому що дозволяє збільшити швидкість проведення нервового імпульсу і є більш ощадливим енергетично. У цьому випадку деполяризуються тільки невеликі ділянки мембрани, виникають менші втрати іонів, отже, клітині доводиться витрачати менше енергії для забезпечення роботи K+,Na+Fнасосів.
§ 35. Швидкість проведення нервового імпульсу
Розподіл потенціалу дії ϕ в залежності від віддалі x і часу t по немієлінизованому нервовому волокну визначається наступним рівнянням, яке отримало назву телеграфного
Рис. 6.8. Поширення збудження в мієлінизованому нервовому волокні.
де D — діаметр волокна; l — товщина мембрани; Сm — ємність; ρa
— питомий опір аксоплазми; ρm — питомий опір мембрани, що різко знижується при збудженні.
Розв’язок цього рівняння має наступний вигляд
ϕ = ϕ0 . exp(–x/λ),
де ϕ0 — потенціал в точці х = 0; λ — стала довжини волокна, що дорівнює
133

При видаленні від точки виникнення збудження на величину λ потенціал зменшується в е раз. Клітині вигідніше мати великі значення λ, тому що при цьому загасання імпульсу відбувається повільніше. Відсутність повного загасання імпульсу пояснюється тим, що кожний новий потенціал дії підсилює сигнал.
Швидкість проведення нервового імпульсу по немієлінизованих нервових волокнах, так само як і стала довжини, пропорційна квадратному кореню з діаметра волокна. Збільшення діаметра сприяє збільшенню λ і швидкості поширення збудження. Цим пояснюється існування гігантських аксонів головоногих молюсків. Швидкість проведення збудження по немієлінизованому волокну діаметром 1 мкм складає тільки 2 м/с, тоді як для волокон діаметром 0,5–1 мм, ця величина вже досягає 20 м/с.
Для мієлінизованих нервових волокон швидкість проведення збудження залежить від довжини міжперехватних ділянок (ділянок між двома перехватами Ранв’є). Замічено, що час, необхідний для передачі сигналу від одного перехвату до іншого, приблизно однаковий і складає біля 0,07 мс. У той же час довжина міжперехватних ділянок пропорційна діаметру волокна. Таким чином, швидкість проведення нервового імпульсу по мієлінизованих волокнах пропорційна діаметру D, а не  . Тому в організмах, що мають мієлінизовані волокна, відпала необхідність значно збільшувати їхній діаметр. Таке волокно діаметром 20 мкм проводить збудження зі швидкістю 120 м/с.
. Тому в організмах, що мають мієлінизовані волокна, відпала необхідність значно збільшувати їхній діаметр. Таке волокно діаметром 20 мкм проводить збудження зі швидкістю 120 м/с.
Вимір швидкості проведення нервового імпульсу часто проводять у медичній практиці: при порушенні мієлінової оболонки швидкість поширення збудження знижується.
Для нервових волокон приблизно однакового діаметра швидкість поширення збудження залежить від величини фактора надійності η, що виражається так:
де ϕ0 — амплітуда потенціалу дії; ϕk — критичний рівень деполяризації, що викликає генерацію потенціалу дії. Звичайно фактор надійності складає 5—6, тобто потенціал дії здатний збудити наступну ділянку мембрани, тому що його величина значно перевищує граничний рівень деполяризації. Дія місцевих анестетиків (наприклад, новокаїну) зводиться до зниження величини ϕ0 і збільшення ϕk за рахунок інактивації натрієвих каналів, що приводить до зниження фактора надійності.
134

Приклади розв’язання задач
Задача 6.1. Внутрішньоклітинна концентрація іонів калію в 20 разів перевищує зовнішню. Потенціал спокою при цьому дорівнює –80 мВ. Обчислити температуру клітини.
Розв’язання. З рівняння Нернста |
знайдемо |
=310 K = 37 °C.
Задача 6.2. Стала довжини немієлінизованого нервового волокна дорівнює 55 мкм. Потенціал у деякій точці цього волокна дорівнює ϕ0. На якій відстані від цієї точки потенціал зменшиться в три рази?
Розв’язання. Використовуючи вираз ϕ = ϕ0ехр(–х/λ), отримаємо
звідки знаходимо
х = λln 3 = 55·ln3 = 60,4 мкм.
Питання тестового контролю
6.1. Рівноважний мембранний потенціал для іонів натрію дорівнює:
а)
б)
в)
г)
д)
6.2. Рівняння Гольдмана — Ходжкіна — Катца має вигляд:
135

а) 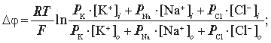
б) 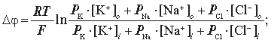
в)
г) 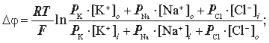
д)
6.3.В спокої потенціал нервової клітини наближується до рівноважного:
а) кальцієвого потенціалу; б) натрієвого потенціалу; в) хлорного потенціалу; г) калієвого потенціалу; д) потенціалу протонів.
6.4.Під час генерації потенціалу дії потенціал нервової клітини наближується до рівноважного:
а) кальцієвого потенціалу; б) натрієвого потенціалу; в) хлорного потенціалу; г) калієвого потенціалу; д) потенціалу протонів.
6.5.Внутрішньоклітинне середовище заряджене в порівнянні з позаклітинним:
а) у спокої негативно, на максимумі потенціалу дії позитивно; б) у спокої позитивно, на максимумі потенціалу дії негативно; в) завжди позитивно; г) завжди негативно.
6.6.Проникність мембрани для іонів калію в спокої:
а) значно більша проникності для іонів натрію; б) значно менша проникності для іонів натрію;
в) приблизно дорівнює проникності для іонів натрію.
6.7. Під час генерації потенціалу дії проникність мембрани для іонів калію:
а) більша проникності для іонів натрію;
136

б) менша проникності для іонів натрію; в) приблизно дорівнює проникності для іонів натрію.
6.8. Сталою довжини нервового волокна називається:
а) відстань, на якій швидкість поширення нервового імпульсу зменшується в е разів;
б) відстань, на якій швидкість поширення нервового імпульсу збільшується в е разів;
в) відстань, на якій швидкість поширення нервового імпульсу зменшується в 10 разів;
г) відстань, на якій швидкість поширення нервового імпульсу збільшується в 10 разів;
д) відстань, на якій величина потенціалу дії збільшується в е разів. 6.9. Якщо в деякій точці немієлінизованого волокна потенціал дорів-
нював ϕ0, то на відстані х від цієї точки він вже буде складати:
а) 
б) 
в) 
г) 
д)  6.10. Швидкість поширення нервового імпульсу:
6.10. Швидкість поширення нервового імпульсу:
а) пропорційна  для немієлінизованих і мієлінизованих нервових волокон;
для немієлінизованих і мієлінизованих нервових волокон;
б) пропорційна D для немієлінизованих і мієлінизованих нервових волокон; 
в) пропорційна  для немієлінизованих і D для мієлінизованих нервових волокон;
для немієлінизованих і D для мієлінизованих нервових волокон; 
г) пропорційна D для немієлінизованих і  для мієлінизованих нервових волокон;
для мієлінизованих нервових волокон;
д) пропорційна D2 для немієлінизованих і мієлінизованих нервових волокон,
де D — діаметр волокна.
137

138
Розділ 7. БІОФІЗИКА М’ЯЗОВОГО СКОРОЧЕННЯ
§ 36. Будова м’язового волокна
Всі м’язи організму поділяються на гладкі і поперечноFсмугасті. Серед останніх розрізняють два типи: кістякові і серцевий. Нижче будуть розглянуті тільки кістякові м’язи.
Мембрана м’язових клітин — сарколема — також, як і мембрана нервових клітин, електрозбудлива і здатна проводити потенціал дії. Ці процеси в м’язових клітинах відбуваються за тим самим принципом, що й у нервових. Потенціал спокою ϕ0 м’язового волокна складає приблизно –90 мВ, що нижчий такого в нервовому волокні (–70 мВ), а критична деполяризація ϕ, по досягненні якої виникає потенціал дії, має однакове значення з нервовим. Тому збудливість м’язового волокна дещо нижча, ніж нервового, тому що клітину потрібно деполяризувати на більш велику величину:
∆ϕм > ∆ϕн, де ∆ϕм =ϕ0м – ϕм, ∆ϕн =ϕ0н – ϕн, індекси “м” і “н” відносяться відповідно до м’язових і нервових клітин.
Відповіддю м’язового волокна на збудження є скорочення, яке робить скорочувальний апарат клітини — міофібрили. Вони являють собою тяжі, що складаються із двох видів ниток: товстих, називаних також міозиновими, і тонких (актинових). Товсті нитки (діаметром 15 нм і довжиною 1,5 мкм) мають у своєму складі тільки один білок — міозин, тонкі (діаметром 7 нм і довжиною 1 мкм) містять три види білків: актин, тропоміозин і тропонін.
Актин являє собою довгу білкову нитку, однак його не можна віднести до фібрилярних білків1 . Він складається з окремих глобулярних білків, зчеплених між собою таким чином, що вся структура являє собою витягнутий ланцюг. Молекули гло- 





булярного актину (GFактину) мають |
Рис. 7.1. Об’єднання окремих глобул |
|
бічні і кінцеві центри зв’язування |
G[актину в F[актин. |
|
|
|
|
1 Поліпептидний ланцюг глобулярних білків укладений в компактну структуру (від лат. globulus — кулька), а у фібрилярних білків витягнута в довгий ланцюг.
139
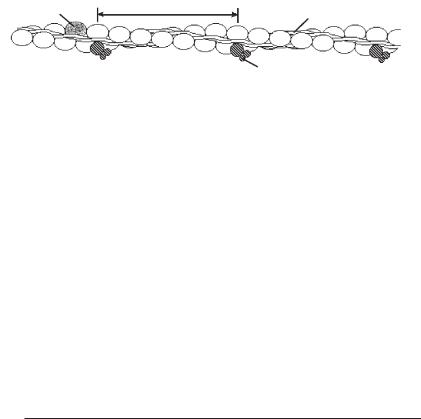
з іншими такими ж молекулами. У результаті вони об’єднуються таким чином, що утворюють структуру, яку часто порівнюють із двома разками, з’єднаних разом (рис. 7.1). Утворена з молекул GFактину стрічка закручена в спіраль. Така структура називається фібрилярним актином (FFактином). Крок спіралі (довжина витка) складає 38 нм, на кожний виток спіралі доводиться 7 пар GFактину. Полімеризація GFактину, тобто утворення FFактину відбувається з використанням енергії АТФ, і навпаки, при руйнації FFактину виділяється АТФ.
Уздовж спіральних борозенок актинових філаментів (ниток) розташовується білок тропоміозин (грец. trope — повертати, mys — м’яз). Кожна нитка тропоміозину, що має довжину 41 нм, складається з двох ідентичних αFланцюгів, разом закручених у спіраль із довжиною витка 7 нм. Уздовж одного витка FFактину розташовані дві молекули тропоміозину. Кожна молекула тропоміозину з’єднується, трохи перекриваючись, із наступною, у результаті тропоміозинова нитка простирається уздовж актину безупинно (рис. 7.2).
У клітинах поперечноFсмугастих м’язів до складу тонких ниток крім
G[актин |
38 нм |
Тропоміозин |
|
Тропонін
Рис. 7.2. Будова тонкої нитки міофібрили.
актину і тропоміозину входить ще і білок тропонін. На кожний крок спіралі FFактину доводиться 2 молекули тропоніну. Цей глобулярний білок має складну будову: він складається з трьох субодиниць, кожна з яких виконує свою функцію в процесі скорочення.
Товста нитка складається з великого числа молекул міозину, зібраних у пучок. Кожна молекула міозину довжиною біля 155 нм і діаметром 2 нм складається із шести поліпептидних ниток: двох довгих і чотирьох коротких (рис. 7.3). Довгі ланцюги разом закручені
в спіраль із кроком 7,5 нм і утворюють фібрилярну частину міозинової |
||
S2 |
|
молекули. На одному із кінців мо- |
|
лекули ці ланцюги розкручуються й |
|
S1 |
|
|
|
утворюють роздвоєний кінець. Кож- |
|
|
|
|
45 нм |
110 нм |
ний з цих кінців утворює комплекс із |
Важкий |
Легкий мероміозин |
двома короткими ланцюгами, тобто |
|
||
|
на кожній молекулі є дві головки. |
|
мероміозин |
|
|
|
|
Це глобулярна частина міозинової |
Рис. 7.3. Будова міозину. |
молекули. У міозині виділяють два |
|
140