

Биофизика 02
.pdfіншу речовину, він повинен, принаймні, мати у своїй структурі шестивуглецевий піранозний кістяк. Багато природних переносників здатні переносити деякі невластиві організму сполуки, наприклад, ліки, якщо ці речовини мають структурну подібність із природним субстратом. Наприклад, протипухлинний препарат із класу азотистих іпритів — сарколізин — містить фенілаланінний залишок і тому здатний транспортуватися в клітину переносником для фенілаланіну.
Явище насичення переносника при високих концентраціях речовин, що транспортуються, можна продемонструвати на прикладі реабсорбції глюкози в ниркових канальцях. З курсу фізіології відомо, що плазма крові видавлюється під дією гідростатичного тиску (явище фільтрації) у нирковий клубочок, а потім реабсорбується назад за винятком продуктів обміну і надлишкових кількостей води та електролітів. Якщо концентрація глюкози в плазмі крові не перевищувала так званий нирковий поріг, величина якого складає 11 ммоль/л (2 г/л), то глюкоза реабсорбується цілком. У противному випадку спостерігається явище насичення системи переносу глюкози і її залишок виділяється із сечею, де в нормі концентрація глюкози повинна практично дорівнювати нулю. Якщо аналізи при цьому показують підвищену концентрацію цукру в крові, то цей факт може свідчити про наявність цукрового діабету в пацієнта. З другого боку, спостерігаються випадки, коли в сечі глюкоза присутня, у той час як її концентрація в крові не підвищена. У цьому випадку, можливо, порушені процеси транспорту глюкози в нирках.
Нирковий поріг реабсорбції спостерігається і для деяких інших речовин, наприклад, фосфату, сульфату, амінокислот. Явище насичення переносників іноді може сприяти підтримці постійної концентрації даної речовини в організмі. У випадку глюкози нирковий поріг удвічі вище її нормальної концентрації в плазмі крові, тому нирки не можуть приймати помітної участі в регуляції кількості глюкози в крові. Для фосфату ж нормальна концентрація в крові майже збігається з величиною ниркового порогу, що приводить до виведення його зайвої кількості із сечею і підтримці його концентрації на постійному рівні.
§ 28. Індукований іонний транспорт
Транспорт іонів за допомогою переносників називається індукованим іонним транспортом, а самий переносник — іонофором. Структура іонофора така, що зовні він покритий неполярними групами, завдяки чому йому вигідно знаходитися в гідрофобній частині мембрани, усередині ж він має порожнину, яка вистелена полярними групами, до яких і приєднуються іони. Багато іонофорів виділені з мікроорганізмів, інші
1Глюкозид, одержаний з кори грушевого дерева.
111

Рис. 5.3. Визначення констант Jmax та
Km за методом Лайнуївера — Берка і вплив конкурентного інгібітора на величину потоку.
1 — потік речовини у відсутність інгібітора; 2 — потік речовини в присутності інгібітора.
мають синтетичне походження. Відповідно до формули Борна (5.15),
енергія, необхідна для переходу іона через мембрану, обернено пропорційна радіусу іона. При зв’язуванні іона з переносником відбувається збільшення радіуса частинки, яка переноситься, що сприяє зниженню енергії переходу.
Пасивний транспорт іонів за допомогою рухливих переносників описується рівнянням, виведеним на основі формули Борна:
д е W — е н е р г і я к о м п л е к с у іонFпереносник; r — радіус іона; b — радіус переносника; εм — діелектрична проникність мембрани; εп — діелектрична проникність внутрішньої сфери переносника. Розрахована по даній формулі енергія комплексу переносника з іонами натрію або калію складає біля 15 кДж/моль, у той час як енергія, необхідна для їхнього переносу через мембрану без участі переносника, складає 250–350 кДж/моль.
Багато переносників є самі по собі нейтральними і, приєднавши до себе іон, пересікають мембрану у вигляді зарядженого комплексу, інші, як, наприклад, нігерицин, іонізовані, а в комплексі з іоном являють собою нейтральні частинки.
Відомі два різновиди іонних переносників. Одні з них, так звані рухливі переносники (рис. 5.4), зв’язуються з іоном, дифундують із ним через мембрану, визволяють його по інший її бік і повертаються назад. При цьому, якщо переносник не залишає мембрану, то говорять, що перенос відбувається за механізмом «малої каруселі», як, наприклад, у випадку переносу іонів валіноміцином, якщо ж переносник виходить із мембрани у міжF або внутрішньоклітинну рідину, де і відбувається зв’язування і вивільнення іона, то — за механізмом «великої каруселі». Інші переносники залишаються нерухомими щодо мембрани, вони формують мембранні канали, що, як правило, не пронизують мембрану наскрізь. Іони, перетинаючи мембрану, змушені переходити по ланцюжку з однієї молекули переносника в іншу, тому такий перенос називається естафетним (рис. 5.5).
Прикладом рухливих переносників є антибіотики валіноміцин і нігерицин. Молекула валіноміцина (рис. 5.6) має циклічну будову. Її внут-
112

рішня порожнина містить гідрофільні групи, із якими здатні зв’язуватися іони. Так як переносник має жорстку будову і розмір його порожнини фіксований, то з валіноміцином здатні зв’язуватися тільки ті іони, що стерично відповідають порожнині переносника. Тому валіноміцин є високоселективним. Основним іоном, який він переносить, є К+. Менший за розмірами іон натрію гірше взаємодіє з групами, що вистилають порожнину каналу, і тому переноситься в 104 рази рідше, ніж К+.
Молекула іншого рухливого переносника — нігерицина являє собою ланцюг із 15 гідрофобних амінокислот, серед яких зустрічаються не тільки LF, але і DFізомери, які ніколи не зустрічаються в природних білках. Молекула нігерицина не замкнута в цикл, а при зв’язуванні з іоном обвиває його навколо. Нігерицин являє собою слабку кислоту (при нейтральних значеннях рН його молекула іонізована) і тому здатний зв’язуватися з протонами й іонами металів. Якщо валіноміцин, відтранспортував К+ на інший бік мембрани, повертається обернено порожнім, то нігерицин здатний переносити іони в обидва кінці при наявності відповідних трансмембранних різниць концентрацій. Наприклад, зв’язавшись із протоном по один бік мембрани й відтранспортувавши його на іншу, нігерицин здатний там зв’язатися з К+ і повернутися разом із ним назад. Цей вид транспорту називається обмінним. Нігерицин та його аналоги не настільки специфічні як нейтральні іонофори.
Одним із найбільше вивчених переносників, що формує канали, є антибіотик граміцидин. Молекула граміцидина являє собою ланцюг, який згорнутий в спіраль таким чином, що утворює канал довжиною 3 нм і внутрішнім діаметром 0,5 — 0,8 нм. Так як такої довжини недостатньо, щоб пронизати мембрану наскрізь, то транспорт може здійснюватися тільки в тому випадку, якщо дві молекули граміцидина утворять послідовний ланцюжок. Граміцидин також не має високої специфічності унаслідок того, що діаметр каналу може змінюватися в залежності від розміру іона.
Багато переносників, що формують канали, наприклад, аламецитін, здатні чергувати відкритий і закритий стан. Причиною цих переходів може бути, наприклад, зміна трансмембранного потенціалу.
§ 29. Активний транспорт
Розглянутий раніше транспорт частинок через мембрану завжди відбувався убік зменшення електрохімічного потенціалу. Однак, в організмі часто виникає потреба транспорту нейтральних частинок або іонів, у процесі якого електрохімічний потенціал зростає. Наприклад, концентрація іонів калію в клітині набагато вища, ніж у міжклітинному середовищі, а натрію навпаки нижча. Високий градієнт концентрації калію підтримується градієнтом електричного потенціалу, направленим
113
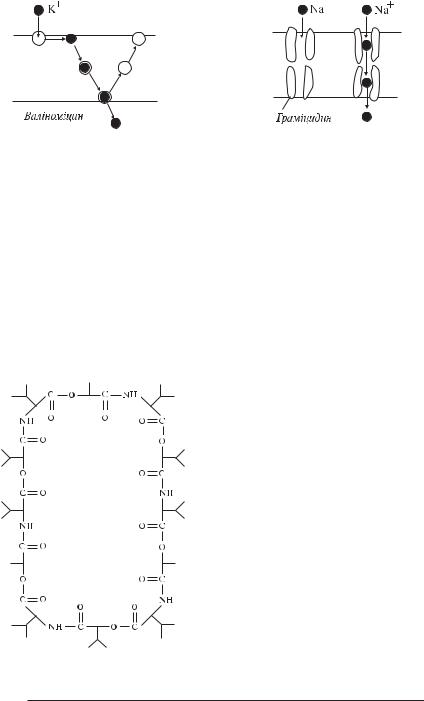
Рис. 5.4. Рухливий переносник. Рис. 5.5. Естафетне перенесення.
у протилежний бік, і підкоряється умові рівноваги відповідно до рівняння Нернста (5.8). Іонам натрію також набагато вигідніше знаходитися в клітині, тому що вони несуть заряд такого ж знака, що й іони калію. Спонтанний винос із клітини іонів натрію неможливий і здійснюється тільки за рахунок притоку енергії. Відомі системи активного транспорту для іонів натрію, калію, кальцію і водню.
Підтримка градієнту концентрацій іонів натрію і калію забезпечується роботою натрійFкалієвого насоса (Na+,K+Fнасоса). Він являє собою мембранні білки, що працюють як ферменти аденозинтрифосфатази (АТФази). Задачею цих ферментів є розщеплення АТФ на АДФ і неорганічний фосфат. Цей процес супроводжується виділенням енергії, що витрачається на транспорт іонів убік збільшення електрохімічного потенціалу. Розщеплення АТФ стимулюється іонами Na+ і K+ і залежить від
наявності Mg2+. Таким чином, активний транспорт можливий тільки за рахунок спряження транспорту якоїсь речовини
з реакцією гідролізу АТФ.
Механізм спряження остаточно не вияснений. Найімовірніше усього енергія АТФ витрачається на зміну
конформації транспортного білка, що змінює його спорідненість (константу зв’язування) до тих або інших іонів.
Транспорт завжди здійснюється в той бік, де спорідненість нижче. У клітині константа зв’язування переносника з Na+ набагато вища, ніж із K+. Тому
іони натрію в клітині зв’язуються з білком і транспортуються в позаклітинне середовище. По інший бік мембрани конформація білка змінюється таким чином, що константа зв’язування з Na+
114

зменшується, а з К+ — збільшується. Структура ділянки білка, яка зв’язує іони, у цьому випадку така, що до неї можуть приєднатися вже не три, а два іони калію, які і переносяться в клітину (рис. 5.7).
Відповідно до сучасних уявлень, процес активного транспорту Na+
іK+ відбувається в сім етапів.
1.У присутності Mg2+ на внутрішньому боці мембрани утворюється комплекс ферменту АТФази з АТФ:
Еi + АТФ → Еi–АТФ,
де Еi — Na+,K+FАТФаза, іонзв’язуючий центр якої знаходиться на внутрішньому боці мембрани; Е—АТФ — комплекс ферменту з АТФ.
2. Приєднання АТФ змінює конформацію ферменту таким чином, що до комплексу, який утворився, приєднуються три іони натрію
Еi–АТФ + 3Na+ → [Еi–АТФ]Na3+.
3. Відбувається фосфорилювання Na+,K+FАТФази і відщеплення АДФ
[Еi–АТФ]Na3+ → [Еi~Р]Na3+ + АДФ.
4. Іонзв’язуючий центр ферменту переміщається відносно товщини мембрани (можливо, унаслідок фліпFфлопFпереходу білка), у результаті чого іон натрію виявляється на зовнішньому боці клітини
[Еi~Р]Na3+ → [Е0~Р]Na3+,
де Е0 — Na+,K+FАТФаза, іонзв’язуючий центр якої знаходиться на зовнішньому боці мембрани.
5. Зовні клітини внаслідок зменшення спорідненості ферменту до іонів натрію і підвищення спорідненості до калію відбувається обмін цими іонами
[Е0~Р]Na3+ + 2К+ → [Е0~Р]К2+ + 3Na+.
6. Фермент із приєднаними іонами калію знову перевертається в мембрані
[Е0~Р]К2+ → [Еi~Р]К2+.
7. Іон калію і неорганічний фосфат визволяються і фермент повертається у початковий стан
[Еi~Р]К2+ → Еi + Р + 2К+.
Таким чином, енергії, яка виділяється при гідролізі однієї молекули АТФ вистачає на те, щоб винести з клітини три іони натрію і внести два іони калію.
Na+,K+Fнасос сприяє підвищенню не тільки градієнтів концентрацій іонів, але і зростанню градієнту електричного потенціалу, тобто є електрогенним, тому що сума внесених зарядів не дорівнює сумі винесених. Міжклітинне середовище набуває більш позитивного заряду в
115

|
|
порівнянні з клітиною за рахунок |
|
|
|
виносу одного “зайвого” позитив- |
|
|
|
ного іона. |
|
|
|
У мембранах саркоплазматич- |
|
|
|
ного ретикулума м’язових клітин |
|
|
|
і цитоплазматичних мембран |
|
|
|
кардіоміоцитів існує Са2+Fнасос, |
|
а |
б |
робота якого багато в чому подібна |
|
з механізмом переносу іонів Na+, |
|||
|
|
||
Рис. 5.7. Дві конформації іонзв’язуючої |
K+Fнасосом. За один цикл, у |
||
порожнини у Na+,K+[насосі: а) усередині |
процесі якого витрачається одна |
||
|
клітини; б) зовні. |
молекула АТФ, переносяться два |
|
|
|
||
іони кальцію.
Активний транспорт протонів може здійснюватися як за допомогою рухливих переносників, наприклад, пластохінона в мембранах хлоропластів, так і через мембранні канали, наприклад, через канали, утвореними молекулами бактеріородопсину в пурпурних мембранах галофільних бактерій. Протонні канали являють собою інтегральні білки, що утворюють внутрішню пору, де містяться ділянки (полярні групи), до яких можуть приєднуватися протони. Енергія АТФ витрачається на зміну конформації білкових молекул, у результаті чого спорідненість одних ділянок зв’язування до протонів знижується, а інших — збільшується, що змушує протон перескочити на ту ділянку каналу, спорідненість якої до протона на даний момент вище. Шляхом таких перескоків з однієї ділянки зв’язування на інший іон і перетинає мембрану.
Перенос Н+ проти градієнтів їхніх концентрацій здійснюється не тільки за рахунок енергії, що виділяється при гідролізі АТФ, але і за рахунок енергії світла. Останній спосіб використовують вищезгадані галофільні бактерії, які на світлі викачують протони з клітини, а енергію градієнту концентрацій, що при цьому створюється, використовують для синтезу АТФ.
Якщо електричні потенціали з обох боків мембрани однакові, то потік іонів, утворений системами активного транспорту, описується рівнянням
(5.18) де c0 — концентрація АТФази, що здійснює перенос іона, у мембрані; Р — коефіцієнт проникності для комплексу іона і білкової молекули; с1 і с2 — концентрації іонів по обидва боки мембрани; К1 і К2 — константи дисоціації комплексу іона і білка по різні боки мембрани.
116

Дане рівняння дозволяє обчислити максимальну різницю концентрацій, яку може створити даний іонний насос. При досягненні стану, коли різниця концентрацій з обох боків мембрани максимальна, потік стане рівним нулю. Тоді з рівняння (5.18) одержимо
Таким чином, чим більше відношення констант дисоціації іона і білка з обох боків мембрани, тим більша різниця концентрацій, яку може створити даний насос. Чим вище К, тим легше білок відз’єднує іони. Тоді концентрація іона буде вища у тій області, де вища константа дисоціації.
Стан рівноваги, при якому потік іонів, що транспортуються активно, дорівнює нулю, у клітині практично недосяжний у зв’язку з тим, що іони, незважаючи на вкрай низьку проникність для них ліпідного бішару, всеFтаки проникають через нього пасивно, тобто в ту область, де їхня концентрація нижче. Функціонування нервових і м’язових клітин вимагає тимчасового відкриття каналів, через які здійснюється пасивне перенесення іонів. Так як концентрація іонів увесь час прагне до вирівнювання, то системи активного іонного транспорту ніколи цілком не припиняють свою роботу, а можуть лише зменшити або збільшити швидкість переносу.
Рівняння (5.18) не враховує існування різниці електричного трансмембранного потенціалу, який зменшує або збільшує швидкість перенесення іона в залежності від знака самого іона і знака потенціалу області, у яку цей іон переноситься.
§30. ВторинноHактивний транспорт
Упопередньому параграфі розглядався транспорт речовин, що проходить зі збільшенням градієнта концентрацій (активний транспорт), джерелом енергії якого служила АТФ. Створені за допомогою систем активного транспорту градієнти концентрацій речовин утворюють на мембрані різницю хімічного або електрохімічного потенціалу, за рахунок чого може здійснюватися перенесення інших речовин проти градієнтів їхніх концентрацій. Такий транспорт, джерелом енергії якого служить не безпосередньо АТФ або енергія окислювальноFвідновлюв альних реакцій, а градієнт концентрацій інших речовин, називається вторинноFактивним або спряженим.
Існують три види вторинноFактивного транспорту іонів: уніпорт, симпорт і антипорт. У першому випадку за рахунок існування на мембрані градієнта електричного потенціалу здійснюється односпрямований транспорт заряджених частинок так, що цей градієнт
117

знижується. Наприклад, мітохондрії в процесі свого функціонування активно викачують протони з матриксу в цитоплазму, у результаті чого їхня внутрішня область виявляється зарядженою більш негативно, ніж зовнішня. У нормальних умовах створений градієнт електрохімічного потенціалу використовується для синтезу АТФ. Якщо ж у клітину додати антибіотик валіноміцин, що є специфічним переносником іонів калію, то починається транспорт цих іонів з цитоплазми у внутрішнє середовище мітохондрій, тобто в область, де електричний потенціал негативний. У даному випадку транспорт К+ хоча і йде проти градієнта концентрацій (у мітохондріях концентрація іонів калію вища, ніж у цитоплазмі), але не вимагає енергії АТФ, тому що здійснюється убік зменшення градієнта електричного потенціалу, створеного різницею концентрацій протонів.
У процесі антипорту (обмінного транспорту або протитранспорту) здійснюється транспорт однаково заряджених іонів двох типів у різні боки. Наприклад, антибіотик нігерицин транспортує протони у той бік, де їх концентрація менша, тобто знижує концентраційний градієнт, але при цьому збільшується градієнт електричного потенціалу. Це, у свою чергу, тягне за собою перенесення іонів калію в протилежний бік (тобто зі зниженням електричного потенціалу), у результаті чого зростає градієнт концентрацій К+.
За механізмом симпорту (котранспорту) здійснюється транспорт протилежно заряджених іонів в один бік. При цьому транспорт одного
зіонів здійснюється пасивно за рахунок наявності градієнта концентрацій, а транспорт другого активно — за рахунок градієнта електричного потенціалу, створюваного транспортом першого іона.
Одним із найбільше вивчених випадків вторинноFактивного транспорту незаряджених молекул є усмоктування глюкози в стінках кишечника. Концентрація глюкози в ентероцитах1 вища, ніж у просвіті кишечника, тому пасивне її усмоктування неможливе. Клітини кишечника активно (за рахунок енергії АТФ) викачують натрій з ентероцитів у серозну область кишечника (тобто туди, де знаходяться кровоносні судини), у результаті чого концентрація Na+ у клітині знижується в порівнянні
зйого концентрацією зовні. Завдяки цьому стає можливим пасивний транспорт Na+ із просвіту кишечника в ентероцит. Однак проста дифузія іонів через бішар украй малоймовірна і Na+ може пересікти мембрану тільки за допомогою переносника. При цьому переносник зв’язується не тільки з Na+, але і з молекулою глюкози. Комплекс, що утворився, пасивно зі зменшенням градієнтів концентрацій іонів Na+ і електричного потенціалу переноситься усередину клітини. Таким чином, транспорт глюкози здійснюється проти градієнта концентрації, але не за рахунок енергії АТФ, а за рахунок існування градієнта концентрації іншої речови-
118
ни, у даному випадку іонів натрію. Подібні системи транспорту існують і для багатьох інших речовин, наприклад, вуглеводів і амінокислот, що дуже важливо, тому що для них відсутні специфічні насоси.
Приклади розв’язання задач
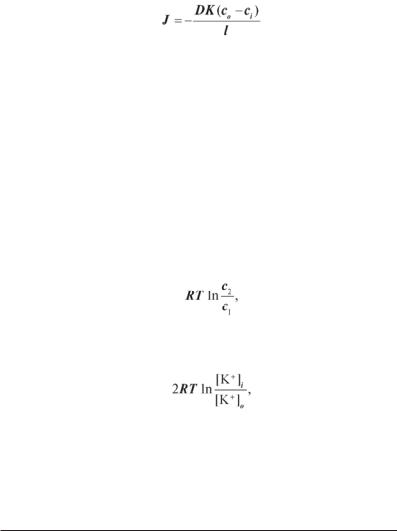
Задача 5.1. Різниця концентрацій іонів на мембрані деякої клітини дорівнює 45 ммоль/л, коефіцієнт розподілу їх між мембраною і навколишнім середовищем дорівнює 30, коефіцієнт дифузії — 1,5·10–8 см2/с, потік — 2,5·10–3 моль/(м2·с). Розрахувати товщину цієї мембрани.
Розв’язання. З рівняння Фіка
одержуємо
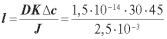 = 8,1·10–9 м = 8,1 нм.
= 8,1·10–9 м = 8,1 нм.
Задача 5.2. Розрахувати енергію, необхідну для здійснення одного циклу Na+,К+FАТФазой у гігантському аксоні кальмара, якщо трансмембранний потенціал цієї клітини складає –60 мВ, концентрація іонів К+ — 0,36 ммоль/м3 і 0,01 ммоль/м3, концентрація іонів Na+ — 0,069 ммоль/м3 і 0,425 ммоль/м3 усередині і зовні, відповідно. Температуру клітини вважати рівною 37 °С.
Розв’язання. Na+,К+Fнасос викачує з клітини три іони натрію і закачує усередину два іони калію, при цьому з клітини виноситься один позитивний заряд. Таким чином, насос робить осмотичну й електричну роботу.
Осмотична робота в загальному вигляді записується як
де с1 — концентрація речовини на початку процесу; с2 — наприкінці. Осмотична робота, необхідна для переносу всередину клітини двох
іонів калію складає:
де [K+]o — концентрація іонів калію зовні клітини (у початковому стані); [K+]i — концентрація іонів калію всередині клітини (у кінцевому стані). Аналогічним способом записується осмотична робота, необхідна для виносу з клітини трьох іонів натрію:
119

де [Na+]i — концентрація іонів натрію всередині клітини (у початковому стані); [Na+]o — концентрація іонів натрію зовні клітини (у кінцевому стані).
Електрична робота дорівнює
z(ϕ2 – ϕ1) = z(ϕ0 – ϕi),
де z=+1, тому що з клітини, яка має потенціал ϕ1 =–60 мВ, викачується один позитивний заряд у середовище з потенціалом ϕ2 = 0 мВ.
Енергія, яку необхідно затратити для вчинення одного циклу Na+,К+FАТФазой, дорівнює сумі цих робіт:
= 18463 + 14050 + 5790 = 38303 Дж/моль ≈ 38 кДж/моль.
Питання тестового контролю
5.1. Коефіцієнт проникності мембрани описується таким виразом:
а) 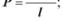 б)
б) 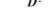 в)
в)  г) д)
г) д)  5.2. Закон Фіка для пасивного транспорту речовин через мембрану
5.2. Закон Фіка для пасивного транспорту речовин через мембрану
має вигляд:
а) 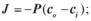 б)
б) 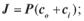 в)
в) 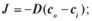 г)
г) 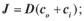
д) 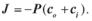
5.3. Рівняння Теорелла має такий вигляд:
а) 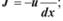
1 Ентероцит — клітина епітелію кишечника.
120