

Вопрос 1 . Анаболические превращения направлены на образование и обновление структурно-функциональных компонентов клетки, т.е. на синтез сложных биомолекул из более простых. Это восстановительные, эндергонические процессы, протекающие с увеличением свободной энергии.
Катаболические превращения направлены на расщепление сложных молекул (как поступивших с пищей, так и уже входящих в состав клеток) до простых компонентов (на конечных стадиях – преимущественно до CO2 и воды). Это окислительные, экзергонические процессы, сопровождающиеся понижением свободной энергии.
Анаболические процессы протекают благодаря энергии, заключенной в химических связях молекул специфической группы «высокоэнергетических» соединений (АТФ и др.), в которых аккумулируется энергия, выделяемая в катаболических процессах. В биохимии под "высокоэнергетическими" соединениями понимаются лабильные вещества, гидролиз которых в физиологических условиях сопровождается значительным понижением ∆G. Выигрыш в свободной энергии используется для смещения равновесия в сопряженных термодинамически невыгодных биохимических процессах, например синтеза биополимеров. Так, АТФ является сопрягающим энергетическим звеном обеих сторон метаболизма – анаболизма и катаболизма. Такое энергетическое сопряжение представляет собой основной способ использования энергии в живых организмах. Но не только АТФ, а и другие соединения, образующиеся в результате катаболизма и используемые в анаболических процессах для синтеза специфических биомолекул, выполняют роль субстратов, сопрягающих отдельные метаболические процессы. Такие пути (или циклы) катаболических и анаболических процессов – называют амфиболические. Амфиболические пути придают обмену веществ значительную гибкость и экономичность с точки зрения использования энергии и материальных ресурсов.
Билет 2. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ
окислительно-восстановительные реакции, протекающие в организмах; осн. источник их энергии. Сводится к передаче атомов водорода или электронов от одного органич. соединения к другому с помощью ферментов-оксидоредуктаз, в первую очередь дегидрогеназ. При анаэробном Б. о. акцептором водорода являются низкомолекулярные соединения (НАД или НАДФ). При аэробном Б. о, (тканевое дыхание) акцептором водорода в конечном итоге служит кислород; ткани при этом поглощают кислород и выделяют CO2 и Н2О. В результате Б. о. освобождённая энергия накапливается в дыхательной цепи в виде химич. связей макроэргич. соединений и используется организмом для различных физиол. функций. Большое значение для Б. о. имеет ЦТК, в результате к-poro окисляются продукты жирового, углеводного и белкового обменов. Энергия освобождается в процессе ферментативного окисления метаболитов специфическими дегидрогеназами. В реакциях дегидрирования электроны и протоны переходят от органических субстратов на коферменты NAD- и FAD-зависимых дегидрогеназ. Электроны, обладающие высоким энергетическим потенциалом, передаются от восстановленных коферментов NADH и FADH2 к кислороду через цепь переносчиков, локализованных во внутренней мембране митохондрий. Восстановление молекулы О2 происходит в результате переноса 4 электронов. При каждом присоединении к кислороду 2 электронов, поступающих к нему по цепи переносчиков, из матрикса поглощаются 2 протона, в результате чего образуется молекула Н2О.
1.Экспериментально было подтверждено, что процесс переноса электронов по ЦПЭ и синтез АТФ энергетически сопряжены. Первый процесс - перенос электронов от восстановленных коферментов NADH и FADH2 через ЦПЭ на кислород - экзергонический. Например:
NADH + Н+ +1/2 O2 → NAD+ + H2O + 52 ккал/моль(≈220 кДж/моль). (1)
Второй процесс - фосфорилирование АДФ, или синтез АТФ, - эндергонический:
АДФ + Н3РО4+7,3 ккал/моль (30,5 кДж/моль) = АТФ + Н2О. (2)
Синтез АТФ из АДФ и Н3РО4 за счёт энергии переноса электронов по ЦПЭ называют окислительным фосфорилированием.
2.Субстратное
S---OP3H2+АДФ→АТФ+S (ФЕРМЕНТ-КИНАЗА)
3.Фотофосфорилирование
H2O→NADH+H++O2 (hv-квант света)
NADH+H++АДФ+P→АТФ+NAD+
Билет 3. Синтез АТФ из АДФ и Н3РО4 за счёт энергии переноса электронов по ЦПЭ называют окислительным фосфорилированием. Отношение количества связанного фосфата к количеству поглощенного кислорода (О) называют коэффициентом фосфорилирования и обозначают как Р/О. Как было показано, коэффициент Р/О равен 3. Эта величина отражает теоретически возможный выход АТФ. В действительности часть энергии электрохимического потенциала используется не на синтез АТФ, а на перенос веществ через митохондриальную мембрану при участии ферментов транслоказ. Суммарный результат окисления (НАДН + Н+) и фосфорилирования АДФ в дыхательной цепи можно представить следующим образом:
НАДН + Н++ ½О2 → НАД+ + Н2О;
3АДФ + 3Н3РО4 → 3АТФ + 3Н2О.
Митохондрии обычно имеют форму цилиндра с закругленными концами, длиной 1–4 мкм и диаметром 0,3–0,7 мкм. Они состоят из внешней и внутренней мембран, которые различаются по составу, свойствам и функциям. Внешняя мембрана легко проницаема для молекул с молекулярной массой до 5 000, в то время как проницаемость внутренней мембраны строго ограничена и избирательна, что определяется наличием специфических транспортных систем. На долю ферментов дыхательной цепи приходится 30–40% всех белков внутренней мембраны.
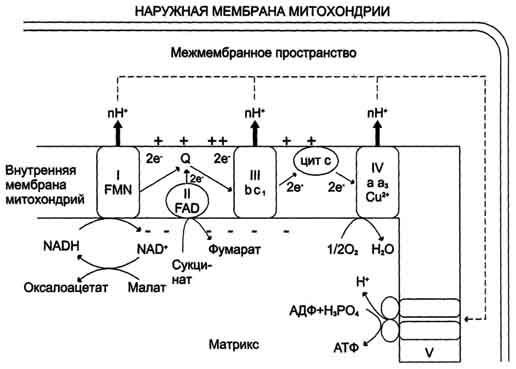
Сопряжение дыхания и синтеза АТФ в митохондриях. I - NADH-дегидрогеназа; II - сукцинатдегидрогеназа; III - QН2-дегидрогеназа; IV - цитохромоксидаза; V - АТФ-синтаза. Энергия протонного потенциала (электрохимического потенциала ΔμН+ используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы.
Билет 4. Дыхательная цепь — система структурно и функционально связанных трансмембранных белков и переносчиков электронов. Она позволяет запасти энергию, выделяющуюся в ходе окисления НАД∙Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану.
Организация дыхательной цепи в митохондриях. Основные переносчики электронов встроены во внутреннюю мембрану митохондрий и организованы в 4 комплекса, расположенных в определённой последовательности. В этой последовательности их стандартные о-в потенциалы становятся более положительными по мере приближения к кислороду. Каждое звено этой цепи специфично в отношении донора и акцептора электронов. На первом этапе дегидрогеназы катализируют отщепление водорода от различных субстратов. Если субстратами служат а-гидрокси-кислоты малат, изоцитрат, 3-гидроксибутират, водород переносится на NAD+. Образовавшийся NADH в дыхательной цепи, в свою очередь, окисляется NADH-дегидрогеназой (комплекс I). Если субстратом служат такие соединения, как сукцинат или глицерол-3-фосфат, акцептором водорода служат FAD-зависимые дегидрогеназы. От NADH и FADH2 электроны и протоны передаются на убихинон и далее через цепь цитохромов к молекулярному кислороду. До сих пор точно неизвестно, каким образом расположены все переносчики электронов дыхательной цепи. Однако установлено, что в расположении дыхательных комплексов существует определённая асимметрия: некоторые из белков-переносчиков находятся ближе к той стороне внутренней мембраны, которая обращена к матриксу, а другие - к противоположной; некоторые белки пронизывают мембрану насквозь.
Комплекс I (НАДН дегидрогеназа) окисляет НАД-Н, отбирая у него два электрона и перенося их на растворимый в липидах убихинон, который внутри мембраны диффундирует к комплексу III. Вместе с этим, комплекс I перекачивает 4 протона из матрикса в межмембранное пространство митохондрии.
Комплекс II (Сукцинатдегидрогеназа) не перекачивает протоны, но обеспечивает вход в цепь дополнительных электронов за счёт окисления сукцината.
Комплекс III (Цитохром bc1 комплекс) переносит электроны с убихинола на два водорастворимых цитохрома с, расположенных на внутренней мембране митохондрии. Убихинол передаёт 2 электрона, а цитохромы за один цикл переносят по одному электрону. При этом туда также переходят 2 протона убихинола и перекачиваются комплексом.
Комплекс IV (Цитохром c оксидаза) катализирует перенос 4 электронов с 4 молекул цитохрома на O2 и перекачивает при этом 4 протона в межмембранное пространство. Комплекс состоит из цитохромов А и А3, которые, помимо гема, содержат ионы меди.
Ингибиторы дыхательной цепи
Некоторые вещества блокируют перенос электронов через комплексы I, II, III, IV .
Ингибиторы I комплекса — барбитураты,ротенон,пиерицидин
Ингибитор II комплекса — малонат.
Ингибитор III комплекса — антимицин А,миксотиазол,стигматтелин
Ингибиторы IV комплекса — сероводород,цианиды,угарный газ, оксид азота.
Билет 5. На первом этапе дегидрогеназы катализируют отщепление водорода от различных субстратов. Если субстратами служат а-гидрокси-кислоты малат, изоцитрат, 3-гидроксибутират, водород переносится на NAD+. Образовавшийся NADH в дыхательной цепи, в свою очередь, окисляется NADH-дегидрогеназой (комплекс I). Если субстратом служат такие соединения, как сукцинат или глицерол-3-фосфат, акцептором водорода служат FAD-зависимые дегидрогеназы. От NADH и FADH2 электроны и протоны передаются на убихинон и далее через цепь цитохромов к молекулярному кислороду. Никотинамидзависимыедегидрогеназы содержат в качестве коферментов NAD+ или NADP+ . NAD+ и NADP+ - производные витамина PP. Эти коферменты входят в состав активных центров дегидрогеназ, но могут обратимо диссоциировать из комплекса с апоферментами и включаются в состав фермента в ходе реакции. Субстраты NAD- и NADP-зависимых дегидрогеназ находятся в матриксе митохондрий и в цитозоле. Рабочей частью никотинамидных коферментов служит никотинамид. Большинство дегидрогеназ, поставляющих электроны в ЦПЭ, содержат NAD+. Они катализируют реакции типа:
R-CHOH-R1 + NAD+↔ R-CO-R1 + NADH + Н+.
Таким образом, NAD+, присоединяя протоны и электроны от различных субстратов, служит главным коллектором энергии окисляемых веществ и главным источником электронов, обладающих высоким энергетическим потенциалом, для ЦПЭ. Молекулы убихинона различаются длиной углеводородной цепи, обозначается как Q10. В процессе переноса электронов с NADH-дегидрогеназы через FeS на убихинон он обратимо превращается в гидрохинон. Убихинон выполняет коллекторную функцию, присоединяя электроны от NADH-дегидрогеназы и других флавинзависимыхдегидрогеназ, в частности, от сукцинат-дегидрогеназы. Убихинон участвует в реакциях типа:
Е (FMNH2) + Q → Е (FMN) + QH2.
QН2-дегидрогеназа (коэнзим Q-цитохром С-редуктаза, комплекс III) состоит из 2 типов цитохромов (b1 и b2) и цитохрома с1. QН2-дегидрогеназа переносит электроны от убихинола на цитохром с. Внутри комплекса III электроны передаются от цитохромов b на FeS-центры, на цитохром с1, а затем на цитохром с. Группы тема, подобно FeS-центрам, переносят только по одному электрону. Таким образом, от молекулы QH2 2 электрона переносятся на 2 молекулы цитохрома b. В качестве промежуточного продукта в этих реакциях переноса электронов возможно образование свободного радикала се-михинона. В цитохромах типа b гем не связан ковалентно с белком, а в цитохромах с1 и с он присоединяется к белку при помощи тиоэфирных связей). Эти связи образуются путём присоединения 2 цистеиновых остатков к винильным группам гема.
Цитохром с - периферический водорастворимый мембранный белок с молекулярной массой 12 500 Д, имеющий одну полипептидную цепь из 100 аминокислотных остатков, и молекулу гема, ковалентно связанную с полипептидом.
Билет 6. Энергия электрохимического потенциала (∆μH+) используется для синтеза АТФ, если протоны возвращаютсяв матрикс через ионные каналы АТФ-синтазы.
Строение АТФ-синтазы и синтез АТФ
АТФ-синтаза (Н+-АТФ-аза) - интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1. Гидрофобный комплекс F0 погружён в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс. Комплекс F1 выступает в митоховдриальный матрикс. Он состоит из 9 субъединиц (Зα, 3β, γ, ε, δ). Субъединицы аир уложены попарно, образуя "головку"; между α- и β-субъединицами располагаются 3 активных центра, в которых происходит синтез АТФ; γ-, ε-, δ- субъединицы связывают комплекс F1 с F0. Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμH+ заставляет протоны двигаться по каналу АТФ-синтазыв матрикс. Параллельно под действием ΔμH+ происходят конформационные изменения в парах α, β-субъединиц белка F1, в результате чего из АДФ и неорганического фосфата образуется АТФ. Электрохимический потенциал, генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используют для синтеза одной молекулы АТФ.
Билет 7. Дыхательный контроль
Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Если АТФ не используется и его концентрация в клетках возрастает, то прекращается и поток электронов к кислороду. С другой стороны, расход АТФ и превращение его в АДФ увеличивает окисление субстратов и поглощение кислорода. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Механизм дыхательного контроля характеризуется высокой точностью и имеет важное значение, так как в результате его действия скорость синтеза АТФ соответствует потребностям клетки в энергии. Запасов АТФ в клетке не существует.
Гипоэнергетические состояния
Все живые клетки постоянно нуждаются в АТФ. Клетки мозга потребляют АТФ для синтеза нейромедиаторов, в печени происходит синтез гликогена, жиров, белков и др, в миокарде постоянно совершается механическая работа, необходимая для циркуляции крови; мышцы потребляют АТФ при нагрузке. Запасов АТФ в клетках практически не существует. для постоянного синтеза АТФ клеткам необходим приток метаболитов как субстратов дыхания и кислорода как конечного акцептора электронов в реакциях окисления, сопряжённых с синтезом АТФ. Нарушения какого-либо этапа метаболизма, приводящие к прекращению синтеза АТФ, гибельны для клетки. Состояния, при которых синтез АТФ снижен, объединяют термином "гипоэнергетические". Причинами гипоэнергетических состояний могут быть голодание, гиповитаминозы В1, РР, В2; гипоксия. Частой причиной гипоэнергетических состояний могут быть нарушения процессов использования кислорода в клетках.Причинами этих нарушений могут быть:
действие ингибиторов и разобщителей в ЦПЭ;
железодефицитные анемии;
снижение уровня гемоглобина и других железосодержащих белков (цитохромов, FeS-белков), в результате чего нарушаются перенос электронов и синтез АТФ;
наследственные дефекты ферментов ЦПЭ и цитратного цикла.
Примерно 13 из 100 белков, участвующих в окислительномфосфорилировании, кодируются митохондриальной ДНК: 7 субъединиц комплекса I, субъединица комплекса III, 3 субъединицы комплекса IV и 2 субъединицы комплекса V, а также необходимые компоненты их трансляции. Остальные митохондриальные белки синтезируются в ядре.
Ядерная ДНК кодирует более 70 субъединиц белков, участвующих в окислительном фосфорилировании. Нарушения окислительного фосфорилирования в основном связаны с мутациями в митохондриальной ДНК, которые случаются примерно в 10 раз чаще, чем в ядерной. Ткани с высокой потребностью в АТФ (ЦНС, скелетные мышцы, миокард, почки и печень) наиболее чувствительны к нарушениям окислительного фосфорилирования. Дефекты митохондриальной ДНК наследуются по материнской линии, так как митохондрии из клеток сперматозоидов не проникают в оплодотворённую яйцеклетку. Мутации митохондриальной ДНК - частая причина, так как митохондрии не имеют такой же эффективной системы репарации ДНК, как ядро. Даже у здоровых индивидуумов соматические мутации снижают с возрастом возможности окислительного фосфорилирования. В этих случаях способность к синтезу АТФ ниже тканеспецифического уровня нормальных клеток.
Билет 8. Разобщение дыхания и фосфорилирования
Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану вматрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это явление называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается. В этом случае скорость окисления NADH и FADH2 возрастает, возрастает и количество поглощённого кислорода, но энергия выделяется в виде теплоты, и коэффициент Р/О резко снижается. Как правило, разобщители - липофильные вещества, легко проходящие через липидный слой мембраны. Одно из таких веществ - 2,4-динитрофенол легко переходящий из ионизированной формы в неионизированную, присоединяя протон в межмембранном пространстве и перенося его в матрикс. Примерами разобщителей могут быть также некоторые лекарства, например дикумарол - антикоагулянт (или метаболиты, которые образуются в организме, билирубин , тироксин. Все эти вещества проявляют разобщающее действие только при их высокой концентрации.
Терморегуляторная функция ЦПЭ
На синтез молекул АТФ расходуется примерно 40-45% всей энергии электронов, переносимых по ЦПЭ, приблизительно 25% тратится на работу по переносу веществ через мембрану. Остальная часть энергии рассеивается в виде теплоты и используется на поддержание температуры тела. Кроме того, дополнительное образование теплоты может происходить при разобщении дыхания и фосфорилирования. Разобщение окислительного фосфорилирования может быть биологически полезным. Оно позволяет генерировать тепло для поддержания температуры тела у новорождённых, у зимнеспящих животных и у всех млекопитающих в процессе адаптации к холоду. У новорождённых существует особая ткань, специализирующаяся на теплопродукции посредством разобщения дыхания и фосфорилирования - бурый жир. Бурый жир содержит много митохондрий. В мембране митохондрий имеется большой избыток дыхательных ферментов по сравнению с АТФ-синтазой. Около 10% всех белков приходится на так называемый разобщающий белок (РБ-1) - термогенин. По своей структуре термогенин близок к АТФ/АДФ-антипортеру, но не способен к транспорту нуклеотидов, хотя сохранил способность переносить анионы жирных кислот, служащих разобщителями. На внешней стороне мембраны анион жирной кислоты присоединяет протон и в таком виде пересекает мембрану; на внутренней стороне мембраны диссоциирует, отдавая протон вматрикс и тем самым снижает протонный градиент. Образующийся анион возвращается на наружную сторону мембраны с помощью АТФ/ АДФ-антипортера. При охлаждении стимулируется освобождение норадреналина из окончаний симпатических нервов. В результате происходят активация липазы в жировой ткани и мобилизация жира из жировых депо .Образующиеся свободные жирные кислоты служат не только "топливом", но и важнейшим регулятором разобщения дыхания и фосфорилирования.
Билет 9
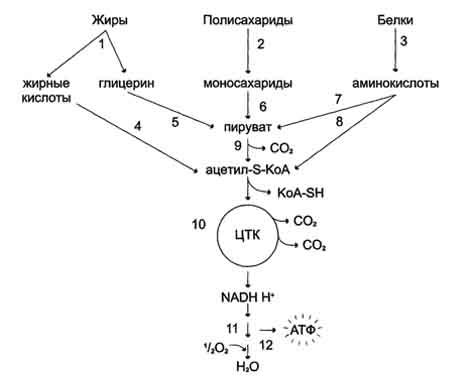
Катаболизм основных пищевых веществ. 1-3 - пищеварение; 4-8 - специфические пути катаболизма; 9-10 - заключительный (общий путь) катаболизма; 11 - ЦПЭ; 12 - окислительное фосфорилирование.
Специфический путь катаболизма - это совокупность превращений, свойственных только 1 соединению.этот путь различен для разных классов соединений.Из глюкозы при гликолизе образуется ПВК. Из жирных кислот при бета-окислении обр-ся Ко-А. Аминокислоты образуют ПВК. В результате специфических путей катаболизма продукты переваривания пищевых веществ (моносахариды, глицерин, жирные кислоты, аминокислоты) превращаются всего в два соединения – пировиноградную кислоту и ацетил-КоА, которые затем направляются в общий путь катаболизма, включающий в себя процесс декарбоксилирования пировиноградной кислоты и цикл трикарбоновых кислот. Некоторые специфические пути включаются в общий путь на стадии пирувата(аниона пировиноградной кислоты), другие – на стадии ацетил-КоА.
Ряд веществ поступает в общий путь катаболизма на промежуточных стадиях цикла трикарбоновых кислот. Именно общий путь катаболизма является источником основной массы первичных доноров водорода для дыхательной цепи. Окислительноед декарбоксилирование пировиноградной кислоты. можно отразить следующим суммарным уравнением:
H3CC(О) COOH+ HS-KoA + НАД+→Н3С-C(O)- S- KoA + НАДН + Н+ + СО2
Пируват Ацетил-КоА
В результате этой реакции образуются ацетил-КоА, восстановленная форма (НАДН + Н+) и диоксид углерода.
Билет 10. Окислительное декарбоксилирование пирувата. происходит в матриксе митохондрий. Транспорт пирувата в митохондриальный матрикс через внутреннюю мембрану митохондрий осуществляется при участии специального белка-переносчика по механизму симпорта с Н+. Превращение пирувата в ацетил-КоА описывают следующим суммарным уравнением:
СН3-СО-СООН + NAD+ + HSKoA → CH3-CO ∼SKoA + NADH + H+ + CO2
В ходе этой реакции происходит окислительное декарбоксилирование пирувата, в результате которого карбоксильная группа удаляется в виде СО2, а ацетильная группа включается в состав ацетил- КоА. Один атом водорода оказывается в составе NADH, а другой в виде Н+ поступает в среду. Реакция необратима, поскольку ΔG0' = -33,5 кДж/моль.
Превращение пирувата в ацетил-КоА включает 5 стадий:
Стадия I. На этой стадии пируват соединяется с ТДФ в составе Е1 и подвергается декарбоксилированию.
Пируват + Е1-ТДФ → Гидроксиэтил-ТДФ + CO2.
В результате этой реакции образуется производное ТДФ с гидроксиэтильной группой при тиазоловом кольце Стадия П. Дигидролипоилтрансацетилаза (Е2) катализирует перенос атома водорода и ацетильной группы от ТДФ на окисленную форму липоиллизиновых групп с образованием ацетилтиоэфира липоевой кислоты
Стадия III. На стадии III КоА взаимодействует с ацетильным производным Е2, в результате чего образуются ацетил-КоА и полностью восстановленный липоильный остаток, простетическая группа Е2
Стадия IV. На стадии IV дигидролипоилде-гидрогеназа (Е3) катализирует перенос атомов водорода от восстановленных липоильных групп на FAD - простетическую группу фермента Е3.
Стадия V . На стадии V восстановленный FADH2 передаёт водород на NAD+ с образованием NADH.
Билет 11. Строение пируватдегидрогеназного комплекса.
Процесс окислительного декарбоксилирования пирувата катализирует сложнооргани-зованный пируватдегидрогеназный комплекс. В пируватдегидрогеназный комплекс (ПДК) входят 3 фермента: пируватдекарбоксилаза (Е1), дигидролипоилтрансацетилаза (Е2) и дигидролипоилдегидрогеназа (Е3), а также 5 коферментов: тиаминдифосфат (ТДФ), липоевая кислота,. FAD, NAD+ и КоА. Кроме того, в состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза
Пируватдегидрогеназный комплекс (ПДК) млекопитающих
|
Фермент |
|
Число мономеров |
Кофермент |
Витамин |
|
1. Пируватдекарбоксилаза (пируватдегидрогеназа) |
E1 |
120 (30 тетрамеров) |
ТДФ |
B1 |
|
2. Дигидролипоилтрансацетилаза |
Е2 |
180(60тримеров) |
Липоамид |
Липоевая кислота (ЛК) |
|
KoA |
Пантотеновая кислота | |||
|
3. Дигидролипоилдегидрогеназа |
Е3 |
12 (6 димеров) |
FAD NAD+ |
В2РР |
Билет12. Цикл лимонной кислоты центральная часть ОПК, циклич аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных промеж продуктовдо CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая участие в синтезеАТФ.это ключевой этапдыхания всех клеток, использующих кислород, Кроме значительной энергетической роли цикл это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются как АК, ЖК углев.
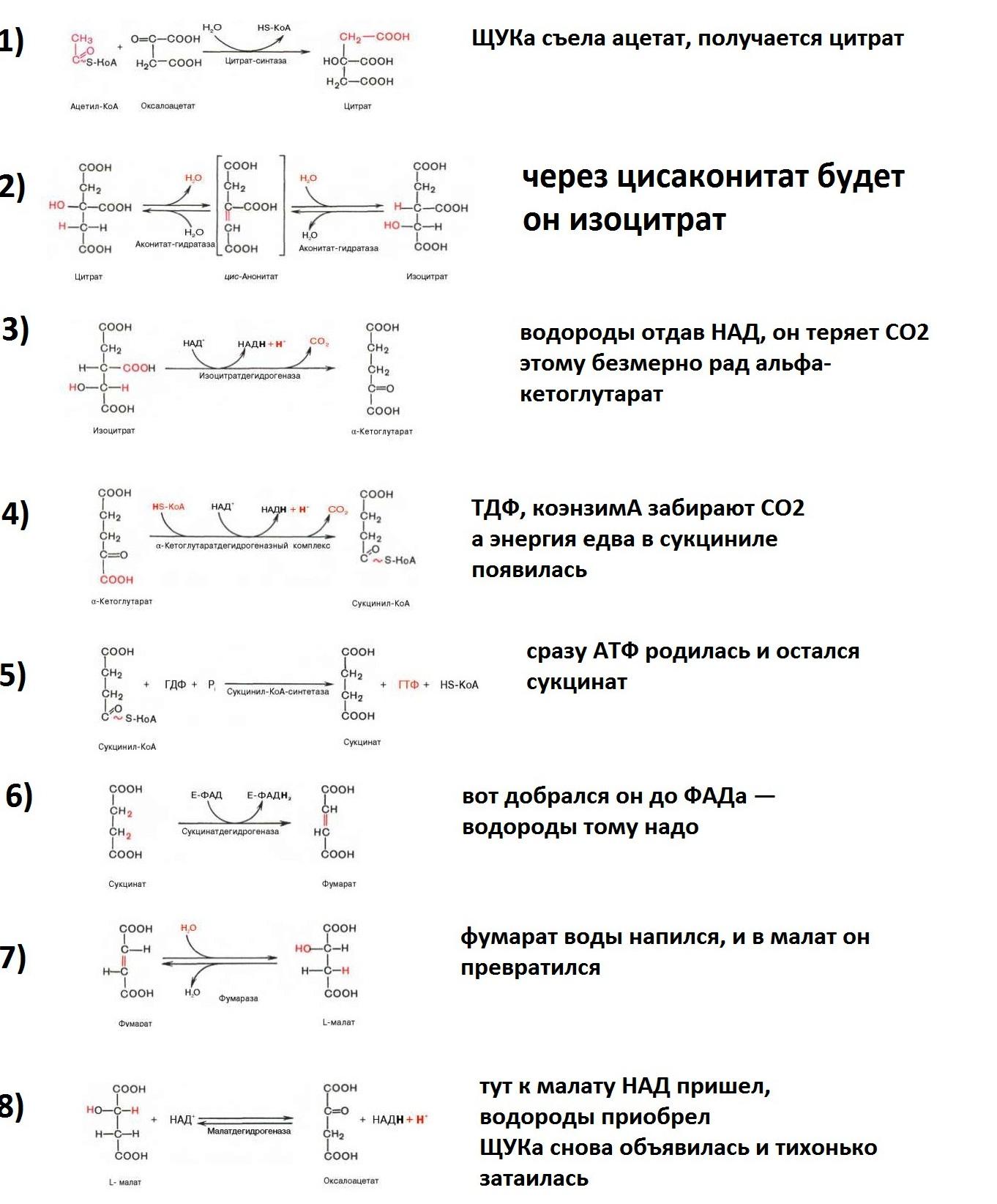
Билет 13. Ключевые реакции цикла лимонной кислоты
Образование цитрата
В реакции образования цитрата углеродный атом метильной труппы ацетил-КоА связывается с карбонильной группой оксалоацетата (одновременно расщепляется тиоэ-фирная связь и освобождается коэнзим A . Реакция сопровождается потерей большого количества энергии в виде теплоты. Катализирует реакцию цитрат синтаза, фермент, локализованный в матриксе митохондрий.
Превращение цитрата в изоцитрат
обратимое превращение цитрата в изоцитрат Фермент, катализирующий эту реакцию, назван аконитазой по промежуточному продукту, цис-аконитовой кислоте, которая предположительно образуется в реакции. Однако это соединение не обнаруживается в свободном виде, так как не отделяется от активного центра фермента до завершения реакции.
Окислительное декарбоксилирование изоцитрата
Эту реакцию катализирует изоцитратдегидрогеназа. Существуют 2 формы изоцитратдегидрогеназы: одна содержит в качестве коферментa
NAD+, вторая - NADP+. NAD-зависимый фермент локализован в митохондриях и участвует в ЦТК; NADP-зависимый фермент, присутствующий и в митохондриях, и В) цитоплазме, играет иную метаболическую роль, В результате действия этого фермента на изоцитрат образуется α-кетоглутарат. Реакция, катализируемая NAD-зависимой изоцитратдегидрогеназой, - самая медленная реакция цитратного цикла. АДФ - аллостерический активатор фермента.
Окислительное декарбоксилирование α-кетоглутарата
α-кетоглутарат подвергается окислительному декарбоксилированию с образованием в качестве конечных продуктов сук-цинил-КоА, СО2 и NADH + Н+. В результате этой реакции образуется сукцинил-КоА. Реакцию катализирует α-кетоглутаратдегидрогеназный комплекс, который по структуре и функциям сходен с пируватдегидрогеназным комплексом (ПДК). Подобно ПДК, он состоит из 3 ферментов: α-кетоглутаратдекарбоксилазы, дигидролипоилтранссукцинилазы и дигидролипоилдегидрогеназы. Кроме того, в этот ферментный комплекс входят 5 коферментов: тиаминдифосфат, кофермент А, липоевая кислота, NAD+ и FAD. она не имеет сложного механизма регуляции, , в этом комплексе отсутствуют регуляторные субъединицы.
Превращение сукцинил-КоА в сукцинат
Сукцинил-КоА - высокоэнергетическое соединение. В митохондриях разрыв тиоэфирной связи сук-цинил-КоА сопряжён с реакцией фосфорилирования (ГДФ) до ГТФ). Эту сопряжённую реакцию катализирует сукцинаттиокиназа. Промежуточный этап реакции - фосфорилирование молекулы фермента по одному из гистидиновых остатков активного центра. Затем остаток фосфорной кислоты присоединяется к ГДФ с образованием ГТФ. С ГТФ концевая фосфатная группа может переноситься на АДФ с образованием АТФ; эту обратимую реакцию катализирует нуклеозид-дифосфаткиназа.
ГТФ + АДФ ↔ ГДФ + АТФ.
Образование высокоэнергетической фосфо-ангидридной связи за счёт энергии субстрата (сукцинил-КоА) - пример субстратного фосфорилирования.
Дегидрирование сукцината
сукцинат превращается в фумарат под действием сукцинатдегидрогеназы. Этот фермент - флавопротеин, молекула которого содержит прочно связанный кофермент FAD. Сукцинат дегидрогеназа прочно связана с внутренней митохондриальной мембраной. Она состоит из 2 субъединиц, одна из которых связана с FAD. Кроме того, обе субъединицы содержат железо-серные центры; одна - Fe2S2, a другая - Fe4S4. В железо-серных центрах атомы железа меняют свою валентность, участвуя в транспорте электронов.
Образование малата из фумарата
Образование малата происходит при участии фермента фумаратгидратазы. Этот фермент более известен как фумараза. Фумараза - олигомерный белок, состоящий из 4 идентичных полипептидных цепей. Он расположен в матриксе митохондрий. Фумаразу относят к ферментам с абсолютной субстратной специфичностью: она катализирует гидратацию только транс-формы фумарата.
Дегидрирование малата
В заключительной стадии цитратного цикла малат дегидрируется с образованием оксалоа-цетата. Реакцию катализирует NAD-зависимая малатдегидрогеназа, содержащаяся в матриксе митохондрий. Равновесие малатдегидрогеназной реакции сильно сдвинуто влево.
Билет14. Анаплеротические реакции цикла лимонной кислоты. Анаболическое значение
ЦТК - один из амфиболических путей метаболизма. В нём осуществляются не только окислительные превращения энергетических субстратов до конечных продуктов СО2 и Н2О, но и происходит образование субстратов для других метаболических путей. Некоторые промежуточные продукты цикла лимонной кислоты: α-кетоглутарат, сукцинат, оксалоацетат могут использоваться для синтеза заменимых аминокислот. Убыль промежуточных продуктов цикла восполняется в реакциях, катализируемых специфическими ферментами. В н.у. реакции, отвлекающие промежуточные продукты из цикла и восполняющие их убыль, находятся в состоянии динамического равновесия, так что концентрация этих продуктов в митохондриях остаётся постоянной. Реакции, обеспечивающие пополнение фонда промежуточных продуктов ЦТК, называются анаплеротическими (пополняющими). Важнейшая из них - реакция синтеза оксалоацетата из пирувата. Эту реакцию катализирует митохондриальный фермент - пируваткарбоксилаза.
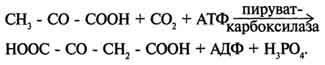
Пируваткарбоксилаза - сложный олигомерный фермент. Молекула фермента содержит 4 простетические группы, представленные биотином , который ковалентно связан амидной связью с е-аминогруппами остатков лизина, находящегося в активном центре фермента. Если для цикла лимонной кислоты не хватает оксалоацетата или какого-нибудь другого промежуточного продукта, то карбоксилирование пирувата ускоряется. В этой реакции в качестве источника энергии используется АТФ. Реакция протекает в 2 стадии.
На первой стадии происходит активация СО2 путём присоединения к одному из атомов азота в молекуле биотина. Эта реакция сопряжена с гидролизом АТФ.
АТФ + СО2+ Е-биотин + Н2О → АДФ + Н3РО4 + Е-биотин-СОО- + 2 Н+.
На второй стадии активированная карбоксильная группа переносится на пируват.
Е-биотин-СОО" + Пируват -" Е-биотин + Оксалоацетат.
Пируваткарбоксилаза - регуляторный фермент. Если концентрация ацетил-КоА увеличивается, то он действует как аллостерический активатор пируваткарбоксилазы, ускоряя образование оксалоацетата. Таким образом, избыток ацетил-КоА способствует активации цитратного цикла. Метаболиты цитратного цикла используются не только как субстраты синтеза углеродного скелета ряда соединений, но и являются донорами водорода для образования восстановленных коферментов, участвующих в реакциях синтеза жирных кислот, стероидов и других веществ ). Два метаболита нитратного цикла могут дегидрироваться при участии NADP-зависимых дегидрогеназ: малата и изоцитрата. Например, малат может поступать из митохондрий в цитозоль клетки. В цитозоле находится NADP-зависимая дегидрогеназа (малик-фермент), катализирующая реакцию:
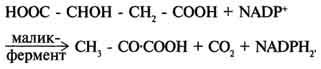
Малат и изоцитрат обеспечивают образование около половины общего фонда NADPH, используемого в восстановительных синтезах; вторая половина образуется в пентозофосфатном пути превращения глюкозы.
Билет 15. Образование активных форм кислорода
ОБРАЗОВАНИЕ ТОКСИЧНЫХ ФОРМ КИСЛОРОДА В ЦПЭ
В ЦПЭ поглощается около 90% поступающего в клетки О2. Остальная часть О2 используется в других окислительно-восстановительных реакциях. Ферменты, участвующие в окислительно-восстановительных реакциях с использованием Кислорода, делятся на 2 группы: оксидазы и оксигеназы. Оксидазы используют молекулярный кислород только в качестве акцептора электронов, восстанавливая его до Н2О или Н2О2.
Оксигеназы включают один (монооксигеназы) или два (диоксигеназы) атома кислорода в образующийся продукт реакции. Хотя эти реакции не сопровождаются синтезом АТФ, они необходимы для многих специфических реакций в обмене аминокислот (см. раздел 9), синтезе жёлчных кислот и стероидов, в реакциях обезвреживания чужеродных веществ в печени. В большинстве реакций с участием молекулярного кислорода его восстановление происходит поэтапно с переносом одного электрона на каждом этапе. При одноэлектронном переносе происходит образование промежуточных высокореактивных форм кислорода. В невозбуждённом состоянии кислород нетоксичен. Образование токсических форм кислорода связано с особенностями его молекулярной структуры. О2 содержит 2 неспаренных электрона с параллельными спинами, которые не могут образовывать термодинамически стабильную пару и располагаются на разных орбиталях. Каждая из этих орбиталей может принять ещё один электрон. Полное восстановление О2 происходит в результате 4 одноэлектронных переходов. Супероксид, пероксид и гидроксильный радикал - активные окислители, что представляет серьёзную опасность для многих структурных компонентов клетки (рис. 6-30). Активные формы кислорода могут отщеплять электроны от многих соединений, превращая их в новые свободные радикалы, инициируя цепные окислительные реакции. Большая часть активных форм кислорода образуется при переносе электронов в ЦПЭ, прежде всего, при функционировании QH2-дегидрогеназного комплекса. Это происходит в результате неферментативного переноса ("утечки") электронов с QH2 на кислород. В отличие от рассмотренного механизма на этапе переноса электронов при участии цитохромоксидазы (комплекс IV) "утечка" электронов не происходит благодаря наличию в ферменте специальных активных центров, содержащих Fe и Сu и восстанавливающих О2 без освобождения промежуточных свободных радикалов. В фагоцитирующих лейкоцитах (гранулоцитах, макрофагах и эозинофилах) в процессе фагоцитоза усиливаются поглощение кислорода и образование активных радикалов. Активные формы кислорода образуются в результате активации NADPH-оксидазы, преимущественно локализованной на наружной стороне плазматической мембраны, инициируя так называемый "респираторный взрыв" с образованием активных форм кислорода. Защита организма от токсического действия активных форм кислорода связана с наличием во всех клетках высокоспецифичных ферментов: супероксиддисмутазы, каталазы, глутатион-пероксидазы, а также с действием антиоксидантов.
Билет 16. Углеводы
СТРОЕНИЕ УГЛЕВОДОВ
Углеводы можно разделить на 3 основные группы в зависимости от количества составляющих их мономеров: моносахариды, олигосахариды и полисахариды.
Моносахариды
Моносахариды - производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от положения в молекуле карбонильной группы моносахариды подразделяют на альдозы и кетозы.
Альдозы содержат функциональную альдегидную группу -НС=О, кетозы содержат кетонную группу >С=О. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, альдогексозы, кетогексозы и т.д. Моносахариды -простым углеводам, так как они не гидролизуются при переваривании, сложные -при гидролизе распадаются с образованием простых углеводов. В пище(фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза. Глюкоза является альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы,, обусловливает химические свойства глюкозы. глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие стереоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. В организме млекопитающих моносахариды находятся в D-конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), аномеры.Фруктоза является кетогексозой (кетогругша находится у второго углеродного атома). Фруктоза так же, как и глюкоза, существует в циклической форме, образуя α- и β-аномеры.
Олигосахариды
Олигосахариды содержат несколько (от двух до десяти) остатков моносахаридов, соединённых гликозидной связью. Дисахариды - наиболее распространённые олигомерные углеводы, встречающиеся в свободной форме. По химической природе дисахариды представляют собой гликозиды, которые содержат 2 моносахарида, соединённые гликозидной связью в α- или β-конфигурации. В пище содержатся в основном такие дисахариды, как сахароза, лактоза и мальтоза. Сахароза- дисахарид, состоящий из α-D-глюкозы и β-D-фруктозы, соединённых α,β-1,2-гликозидной связью. В сахарозе обе аномерные ОН-группы остатков глюкозы и фруктозы участвуют в образовании гликозидной связи. Не относится к восстанавливающим сахарам. Сахароза - растворимый дисахарид со сладким вкусом. Источники-сахарная свёкла, сахарный тростник.Лактоза -молочный сахар; важнейший дисахарид молока млекопитающих. В лактозе аномерная ОН-группа первого углеродного атома остатка D-галактозы связана β-гликозидной связью с четвёртым углеродным атомом D-глюкозы (β-1,4-связь). Поскольку аномерный атом углерода остатка глюкозы не участвует в образовании гликозидной связи,лактоза относится к восстанавливающим сахарам.Мальтозапоступает с продуктами, содержащими частично гидролизованный крахмал, например, солод, пиво. Мальтоза также образуется при расщеплении крахмала в кишечнике. Мальтоза состоит из двух остатков D-глюкозы, соединённых α-1,4-гликозидной связью.Изомальтоза- промежуточный продукт, образующийся при расщеплении крахмала в кишечнике. Состоит из двух остатков D-глюкозы, но соединены эти моносахариды α-1,6-гликозидной связью.
Полисахариды
Структурные различия между полисахаридами определяются:
строением моносахаридов, составляющих цепь;
типом гликозидных связей, соединяющих мономеры в цепи;
последовательностью остатков моносахаридов в цепи.
В зависимости от строения остатков моносахаридов полисахариды можно разделить на гомополисахариды (все мономеры идентичны) и гетерополисахариды (мономеры различны). В зависимости от выполняемых ими функций полисахариды можно разделить на 3 основные группы:
резервные полисахариды, выполняющие энергетическую функцию. Эти полисахариды служат источником глюкозы, используемым организмом по мере необходимости. Резервная функция этих углеводов обеспечивается их полимерной природой. Полисахариды менее растворимы, чем моносахариды, они не влияют на осмотическое давление и поэтому могут накапливаться в клетке, например, крахмал - в клетках растений, гликоген - в клетках животных;
структурные полисахариды, обеспечивающие клеткам и органам механическую прочность полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, Полисахариды межклеточного матрикса водорастворимы и сильно гидратированы.
Билет 17. Моносахариды- производные многоатомных спиртов, содержащие карбонильную группу. В зависимости от положения в молекуле карбонильной группы моносахариды подразделяют на альдозы и кетозы. Альдозы содержат функциональную альдегидную группу -НС=О, кетозы содержат кетонную группу >С=О. Название моносахарида зависит от числа составляющих его углеродных атомов, например альдотриозы, кетотриозы, альдогексозы, кетогексозы и т.д.Моносахариды - простые углеводы, так как они не гидролизуются при переваривании, сложные -при гидролизе распадаются с образованием простых углеводов. В пище (фрукты, мёд, соки) содержится небольшое количество моносахаридов, в основном глюкоза и фруктоза. Глюкоза является альдогексозой. Она может существовать в линейной и циклической формах. Циклическая форма глюкозы, обусловливает химические свойства глюкозы. глюкоза имеет 4 асимметричных углеродных атома, обусловливающих наличие стереоизомеров. Возможно образование 16 стереоизомеров, наиболее важные из которых D- и L-глюкоза. В организме млекопитающих моносахариды находятся в D-конфигурации, так как к этой форме глюкозы специфичны ферменты, катализирующие её превращения. В растворе при образовании циклической формы моносахарида образуются ещё 2 изомера (α- и β-изомеры), аномеры. Фруктоза является кетогексозой (кетогругша находится у второго углеродного атома). Фруктоза так же, как и глюкоза, существует в циклической форме, образуя α- и β-аномеры.
Реакции моносахаридов
Присутствие гидроксильных, альдегидных и кетонных групп позволяет моносахаридам вступать в реакции, характерные для спиртов, альдегидов или кетонов.
Мутаротация, или аномеризация - взаимопревращение аномерных форм моносахаридов, α- и β-формы аномеров находятся в растворе в состоянии равновесия. При достижении этого равновесия происходит мутаротация - размыкание и замыкание пиранового кольца и, соответственно, изменение расположения Н- и ОН-групп при первом углероде моносахарида.
Образование гликозидов. Гликозидная связь имеет важное биологическое значение, потому что именно с помощью этой связи осуществляется ковалентное связывание моносахаридов в составе олиго- и полисахаридов. При образовании гликозидной связи аномерная ОН-группа одного моносахарида взаимодействует с ОН-группой другого моносахарида или спирта. При этом происходят отщепление молекулы воды и образование О-гликозидной связи. Все линейные олигомеры (кроме дисахаридов) или полимеры содержат мономерные остатки, участвующие в образовании двух гликозидных связей, кроме концевых остатков, образующих только одну гликозидную связь. Некоторые гликозидные остатки могут образовывать три гликозидные связи, что характерно для разветвлённых олиго- и полисахаридов. Олиго- и полисахариды могут иметь концевой остаток моносахарида со свободной аномерной ОН-группой, не использованной при образовании гликозидной связи. В этом случае при размыкании цикла возможно образование свободной карбонильной группы, способной окисляться. Такие олиго- и полисахариды обладают восстанавливающими свойствами и поэтому называются восстанавливающими. Аномерная ОН-группа моносахарида может взаимодействовать с NН2-группой других соединений, что приводит к образованию N-гликозидной связи. Подобная связь присутствует в нуклеотидах и гликопротеинах.
Этерификация. Это реакция образования эфирной связи между ОН-группами моносахаридов и различными кислотами. В метаболизме углеводов важную роль играют фосфоэфиры - эфиры моносахаридов и фосфорной кислоты. В метаболизме глюкозы особое место занимает глюкозо-6-фосфат. Образование глюкозо-6-фосфата происходит в ходе АТФ-зависимой реакции при участии ферментов, относящихся к группе киназ. АТФ в данной реакции выступает как донор фосфатной группы. Фосфоэфиры моносахаридов могут образовываться и без использования АТФ. Например, глюкозо-1-фосфат образуется из гликогена при участии Н3РО4. Физиологическое значение фосфоэфиров моносахаридов заключается в том, что они представляют собой метаболически активные структуры. Реакция фосфорилирования моносахаридов важна для метаболизма ещё и потому, что клеточная мембрана мало проницаема для этих соединений, т.е. клетка удерживает моносахариды благодаря тому, что они находятся в фосфорилированной форме.
Окисление и восстановление. При окислении концевых групп глюкозы -СНО и -СН2ОН образуются 3 различных производных. При окислении группы -СНО образуется глюконовая кислота. Если окислению подвергается концевая группа -СН2ОН, образуется глюкуроновая кислота. А если окисляются обе концевые группы, то образуется сахарная кислота, содержащая 2 карбоксильные группы. Восстановление первого углерода приводит к образованию сахароспирта - сорбитола.
Билет18. Дисахариды .
Дисахариды - наиболее распространённые олигомерные углеводы, встречающиеся в свободной форме. По химической природе дисахариды представляют собой гликозиды, которые содержат 2 моносахарида, соединённые гликозидной связью в α- или β-конфигурации. В пище содержатся в основном такие дисахариды, как сахароза, лактоза и мальтоза
Сахароза - дисахарид, состоящий из α-D-глюкозы и β-D-фруктозы, соединённых α,β-1,2-гликозидной связью. В сахарозе обе аномерные ОН-группы остатков глюкозы и фруктозы участвуют в образовании гликозидной связи. не относится к восстанавливающим сахарам. Сахароза - растворимый дисахарид со сладким вкусом. Источники - сахарная свёкла, сахарный тростник.
Лактоза - молочный сахар; важнейший дисахарид молока млекопитающих. В лактозе аномерная ОН-группа первого углеродного атома остатка D-галактозы связана β-гликозидной связью с четвёртым углеродным атомом D-глюкозы (β-1,4-связь). Поскольку аномерный атом углерода остатка глюкозы не участвует в образовании гликозидной связи,лактоза относится к восстанавливающим сахарам.
Мальтоза поступает с продуктами, содержащими частично гидролизованный крахмал, например, солод, пиво. Мальтоза также образуется при расщеплении крахмала в кишечнике. Мальтоза состоит из двух остатков D-глюкозы, соединённых α-1,4-гликозидной связью.
Изомальтоза - промежуточный продукт, образующийся при расщеплении крахмала в кишечнике. Состоит из двух остатков D-глюкозы, но соединены эти моносахариды α-1,6-гликозидной связью
Билет 19. Полисахариды— класс сложных высокомолекулярных углеводов, молекулы которых состоят из десятков, сотен или тысяч мономеров — моносахаридов. Полисахариды необходимы для жизнедеятельности животных и растительных организмов. Они являются одним из основных источников энергии, образующейся в результате обмена веществ организма. Они принимают участие в иммунных процессах, обеспечивают сцепление клеток в тканях, являются основной массой органического вещества в биосфере.
Крахмал:Крахмал состоит из 2 полисахаридов - амилозы и амилопектина, образованных остатками глюкозы. Химическая формула крахмала (C6H10O5)n. Служит основным поставщиком углеводов — одного из важнейших компонентов пищи. Под действием ферментов крахмал гидролизуется до глюкозы, которая окисляется в клетках до углекислого газа и воды с выделением энергии, необходимой для функционирования живого организма.
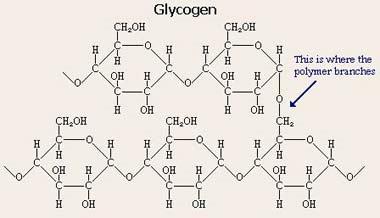
Гликоген: Гликоген — (C6H10O5)n, полисахарид, образованный остатками глюкозы, связанными α-1→4 связями (α-1→6 в местах разветвления); основной запасной углевод человека и животных.
Гетерополисахариды( на картинке 3 представителя, они подписаны) :
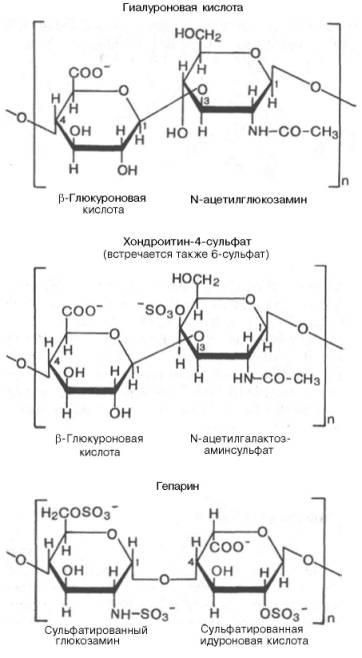
Гетерополисахариды играют важную роль в жизнедеятельности животных организмов, в которых они являются основой углеводных компонентов соединительных тканей (например, хрящей), входят в состав костей и различных слизей. Особенно значительную роль гетерополисахариды играют во многих сложных биохимических системах. Гетерополисахариды составляют основу так называемых гликопептидов — сложных комплексов белков с углеводами, широко представленных в различных тканях организмов. Гетерополисахариды, содержащие аминосахара (а иногда и полипептидиые цепи), играют важную роль в вопросах иммунитета, обеспечивая специфичность иммунологических реакций. Этого же типа гетерополисахариды определяют групповую принадлежность крови.
Билет 20.Переваривание углеводов:


Транспорт моносахаридов в клетки слизистой оболочки кишечника может осуществляться разными способами: путём облегчённой диффузии и активного транспорта. В случае активного транспорта глюкоза и Na+ проходят через мембраны с люминальной стороны, связываясь с разными участками белка-переносчика. При этом Na+ поступает в клетку по градиенту концентрации, и одновременно глюкоза транспортируется против градиента концентрации. Следовательно, чем больше градиент Na+, тем больше поступление глюкозы в энтероциты. Если концентрация Na+ во внеклеточной жидкости уменьшается, транспорт глюкозы снижается. Глюкозные транспортёры (ГЛЮТ) обнаружены во всех тканях.
ГЛЮТ-1 обеспечивает стабильный поток глюкозы в мозг;
ГЛЮТ-2 обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов и печени. ГЛЮТ-2 участвует в транспорте глюкозы в β-клетки поджелудочной железы;
ГЛЮТ-3 обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей;
ГЛЮТ-4 - главный переносчик глюкозы в клетки мышц и жировой ткани;
ГЛЮТ-5 встречается, главным образом, в клетках тонкого кишечника. Его функции известны недостаточно.
Нарушения функции ГЛЮТ-4 возможны на следующих этапах: передача сигнала инсулина о перемещении этого транспортёра к мембране; перемещение транспортёра в цитоплазме; включение в состав мембраны; отшнуровывание от мембраны и т.д.
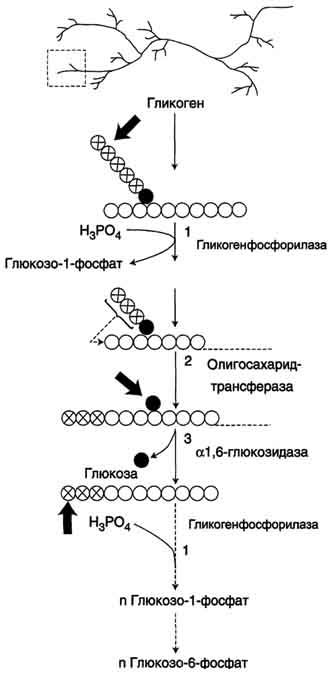
Билет 21.Синтез (слева) и распад(справа) гликогена:
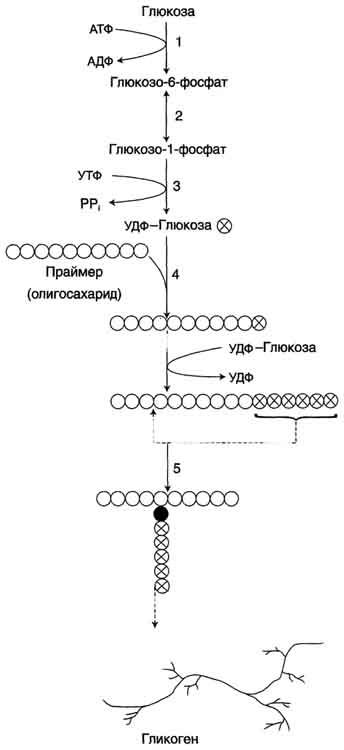
Синтез гликогена. 1 - глюкокиназа или гексокиназа; 2 - фосфоглюкомутаза; 3 - УДФ-глюкрпирофосфорилаза; 4 - гликогенсинтаза (глюкозилтрансфераза); 5 - фермент "ветвления" (амило-1,4 → 1,6-глюкозилтрансфераза),
Распад гликогена. 1 - Гликогенфосфорилаза; 2 - олигосахаридтрансфераза; 3 - α-1,6-глюкозидаза.
Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи). синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т.е. требующим затрат энергии.Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида, называемой "затравка", или "праймер". Распад гликогена или его мобилизация происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени распадается в основном в интервалах между приёмами пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической работы. Распад гликогена происходит путём последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Гликозидная связь расщепляется с использованием неорганического фосфата, поэтому процесс называется фосфоролизом, а фермент гликогенфосфорилазой. Так же как и синтез, расщепление гликогена начинается с нередуцирующего конца полисахаридной цепи. При этом наличие разветвлённой структуры гликогена облегчает быстрое высвобождение глюкозных остатков, так как чем больше концов имеет молекула гликогена, тем больше молекул гликогенфосфорилазы могут действовать одновременно.
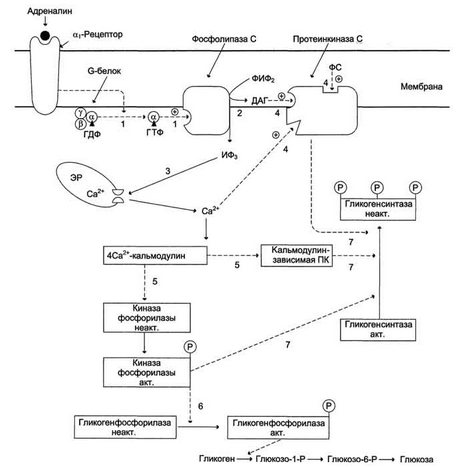
Билет 22. Регуляция синтеза и распада гликогена в печени.
Рисунок 7-29. Рисунок 7-30.
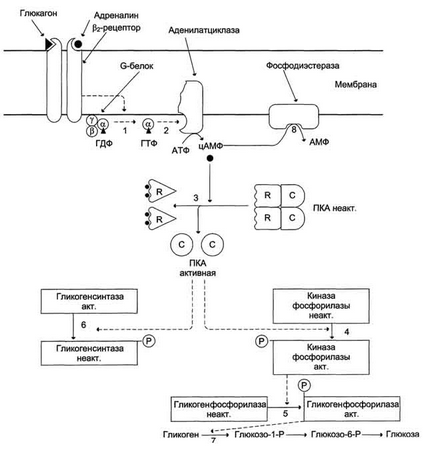


Глюкагон для гепатоцитов служит внешним сигналом о необходимости выделения в кровь глюкозы за счёт распада гликогена (гликогенолиза) или синтеза глюкозы из других веществ – глюконеогенеза.
Адреналин стимулирует выведение глюкозы из печени в кровь, для того чтобы снабдить ткани (в основном мозг и мышцы) "топливом" в экстремальной ситуации. Эффект адреналина в печени обусловлен фосфорилированием (и активацией) гликогенфосфорилазы.
В период пищеварения преобладает влияние инсулина, так как инсулинглюкагоновый индекс в этом случае повышается. В целом инсулин влияет на обмен гликогена противоположно глюкагону. Инсулин снижает концентрацию глюкозы в крови в период пищеварения, действуя на метаболизм печени следующим образом: снижает уровень цАМФ в клетках, фосфорилируя (опосредованно через Ras-путь) и тем самым активируя протеинкиназу В. Активирует (через Ras-путь) фосфопротеинфосфатазу гранул гликогена, которая дефосфорилирует гликогенсинтазу и таким образом её активирует. Индуцирует синтез глюкокиназы, тем самым ускоряя фосфорилирование глюкозы в клетке.
Билет 23. Гликолиз и глюконеогенез.
Глюконеогенез - процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника.
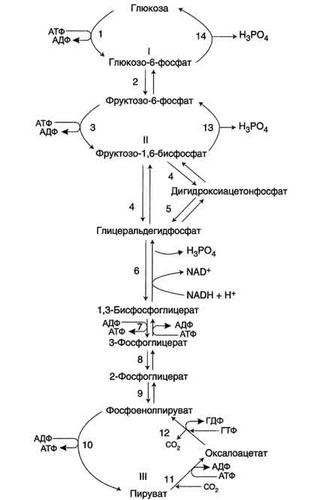
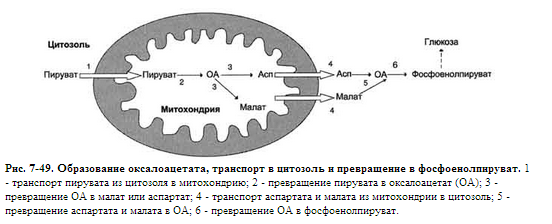
Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 - фосфоглюкоизомераза; 4 - альдолаза; 5 - триозофосфатизомераза; 6 - глицеральдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 - фосфоглицератмутаза; 9 - енолаза. Ферменты необратимых реакций глюконеогенеза: 11 - пируваткарбоксилаза; 12 - фосфоенолпируваткарбоксикиназа; 13 - фруктозо-1,6-бисфосфатаза; 14 -глюкозо-6-фосфатаза. I-III -субстратные циклы. Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза (реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. Однако 3 реакции гликолиза термодинамически необратимы.