

Углеводы — первичные продукты фотосинтеза и основные исходные продукты биосинтеза других веществ в растениях. Составляют существенную часть пищевого рациона. Обмен углеводов — совокупность процессов превращений углеводов в организме. Углеводы, поступающие в организм с пищевыми продуктами, представлены, главным образом, крахмалом и тростниковым сахаром. Крахмал — полисахарид растений, он состоит из цепочки соединенных между собой более простых молекул — моносахаров, главным из которых является глюкоза. По своей структуре крахмал подобен гликогену. Тростниковый сахар — это углевод, который преобладает в нашем рационе. По структуре это дисахарид, т.е. он состоит из двух молекул моносахаров — глюкозы и фруктозы. Глюкоза и фруктоза могут находиться в разных пищевых продуктах и в свободном виде, например, в меде и фруктах. В молочных продуктах содержится, в основном, такой углевод, как лактоза. В организме имеется «депо» углеводов — гликоген, образованный из молекул глюкозы.
Углеводы, прежде всего, источник энергии, в меньшей степени они выполняют
пластическую функцию. Организм человека не нуждается в определенных углеводах. Единственным «незаменимым» производным углеводов, которое обязательно должно поступать с пищей, является аскорбиновая кислота или витамин С, так как у человека отсутствует один из ферментов, необходимых для его синтеза. В сбалансированной диете примерно 50% необходимой человеку энергии должно поступать с углеводами Превращения углеводов связанные с дыхание и брожением
Процесс превращения углеводов начинается с переваривания их в ротовой полости под влиянием слюны, затем некоторое время продолжается в желудке и заканчивается в тонком кишечнике — основном месте гидролиза углеводов под влиянием ферментов, содержащихся в пищеварительном соке поджелудочной железы и тонкого кишечника. Продукты гидролиза — моносахара — всасываются в кишечнике и поступают в кровь воротной вены, по которой моносахариды пищи поступают в печень, где они превращаются в глюкозу. Глюкоза далее поступает в кровь и может вступить в процессы, протекающие в клетках или переходит в гликоген печени.
Роль печени в углеводном обмене
Печень — главный орган, в клетках которого происходят биохимические превращения продуктов пищеварительного гидролиза углеводов и превращение их в глюкозу — форму, доступную для клеток организма. Печень — депо углеводов, так как часть глюкозы хранится здесь в виде гликогена. Печень поддерживает содержание глюкозы в крови на постоянном уровне — в этом состоит глюкостатическая функция печени. При избытке глюкозы в печени происходит синтез гликогена из глюкозы — гликогенез. После приема пищи богатой углеводами содержание гликогена может составлять до 8% веса печени. В среднем, запасы гликогена составляют около 5% веса печени, что у взрослого человека эквивалентно примерно 90 г глюкозы.
При повышении потребности организма в глюкозе происходит распад гликогена печени — гликогенолиз, который достаточен для удовлетворения нужд организма в первые 12-24 часа после приема пищи. Печень — один из главных органов, где происходит процесс ферментативного синтеза глюкозы из углеводных и неуглеводных продуктов — глюконеогенез. Причем клетки печени способны реагировать на возникновение потребности в глюкозе и в клетках других органов. При голодании, после истощения запасов гликогена, процессы глюконеогенеза идут с максимальной интенсивностью, поддерживая «сахар» крови на постоянном уровне. В печени также происходит гликолиз — ферментативный распад глюкозы с освобождением энергии, заключенной в ее молекуле и переводом ее в форму, доступную для организма — т.е. в аденозинтрифосфат (АТФ).
Превращение глюкозы в клетках
В клетках глюкоза может расщепляться как анаэробно (без участия кислорода), так и аэробно (с участием кислорода). В анаэробных условиях гликолиза из каждой молекулы расщепившейся глюкозы образуются 2 молекулы аденозинтрифосфата (АТФ) и 2 молекулы молочной кислоты. При аэробном гликолизе промежуточные продукты углеводного обмена, образующиеся в процессе анаэробного распада углеводов (пировиноградная кислота), не восстанавливаются до молочной кислоты, а окисляются в митохондриях в цикле трикарбоновых кислот до углекислого газа и воды с накоплением энергии в виде АТФ. Кроме того, промежуточные продукты гликолиза являются материалом для синтеза многих важных соединений и используются организмом как еще один источник материала для процессов ассимиляции.
Регуляция содержания глюкозы в крови
О состоянии обмена углеводов можно судить по содержанию сахара в крови. У здорового человека в крови поддерживается постоянная концентрация глюкозы 70-120 мг%. После приема пищи, содержащей углеводы, концентрация глюкозы в крови возрастает примерно до 150мг % и остается на этом уровне около 2 часов, а затем возвращается к норме. Содержание глюкозы в крови — одна из самых важных констант жидкой внутренней среды организма. Ведущая роль в поддержании этой константы на постоянном уровне благодаря идущим там процессам гликогенеза и гликогенолиза принадлежит печени. Длительное повышение содержания глюкозы в крови — гипергликемия стимулирует выделение в кровь инсулина. Инсулин снижаетсодержание глюкозы в крови после возрастания ее концентрации (гипергликемии).
У здорового человека в период между приемами пищи нормальное содержание глюкозы в крови поддерживается путем распада гликогена в печени с образованием свободной глюкозы — процессом гликогенолиза. При снижении сахара крови — гипогликемии, длящейся более длительное время, в кровь поступает глюкагон — гормон, выделяемыйподжелудочной железой. Инсулин, гормон поджелудочной железы, стимулирует процессы синтеза гликогена в печени — гликогенез, поглощение глюкозы клетками других тканей организма, подавляет образование глюкозы, т.е. процессы глюконеогенеза. Инсулин — главный гормон. Этот гормон обладает специфическим действием: он действует исключительно на процессы гликогенолиза, ускоряя образование глюкозы.
При голодании, длящемся более 24 часов, запасы гликогена в печени истощаются. В прессы регуляции включаются гормоны коры надпочечника — глюкокортикоиды. Глюкокортикоиды, во-первых, усиливают глюконеогенез в печени; во-вторых, обеспечивает процессы глюконеогенеза субстратом, усиливая распад белков в тканях организма, они предоставляют для глюконеогенеза углеродсодержащий субстрат. К гормонам, которые обеспечивают повышение сахара крови, относятся адреналин и гормон роста.
Адреналин — гормон мозгового вещества надпочечника. Он усиливает процессы перехода гликогена в глюкозу. Гормон роста, во-первых, подавляет использование глюкозы клетками тканей; во-вторых, при резком и длительном снижении сахара крови стимулирует распад жиров и образование из них углеводов. Дыхание. Все живые организмы дышат, т. е. поглощают кислород и выделяют углекислый газ и воду. При этом происходит разложение органических веществ и выделение энергии, необходимой для жизни каждой клетки, всего растения. В действительности этот процесс многоступенчатый. Он состоит из целого ряда последовательно идущих окислительно-восста-новительных реакций. В качестве органических веществ, необходимых для дыхания, служат в основном углеводы, белки и жиры. Типичным соединением, окисляемым в процессе дыхания, является глюкоза. Энергетически наиболее выгодным для дыхания веществом является жир. 1 г жира при окислении до СО2 и Н2О дает 9,2 ккал, белки — 5,7 ккал, углеводы — 4 ккал. Процесс превращения исходного органического вещества до более простых и затем до СО2 и Н2О требует большого числа различных ферментов.
В
процессе фотосинтеза растения синтезируют
углеводы, которые транспортируются
из листьев в другие органы. На свету и
в темноте клетки растения «дышат»,
окисляя углеводы молекулярным кислородом
с образованием СО2 и
воды. При этом освобождается большое
количество свободной энергии:
С6Н12О6 +
6О2 =
6СО2 +
6Н2О
+ энергия;
∆G![]() =
-2882 кДж/моль (-686 ккал/моль)
Эта
формула в общем виде отражает чрезвычайно
сложный, а главное, контролируемый
процесс, который условно можно разбить
на три этапа: гликолиз,
цикл трикарбоновых кислот и окислительное
фосфорилирование в дыхательной цепи (рис.
1).
=
-2882 кДж/моль (-686 ккал/моль)
Эта
формула в общем виде отражает чрезвычайно
сложный, а главное, контролируемый
процесс, который условно можно разбить
на три этапа: гликолиз,
цикл трикарбоновых кислот и окислительное
фосфорилирование в дыхательной цепи (рис.
1).
Гликолиз и цикл трикарбоновых кислот — это биохимические пути окисления глюкозы, протекающие соответственно в цитозоле и матриксе митохондрий. В биохимических реакциях синтезируется небольшое количество АТФ, и главный их результат — образование соединений с высоким восстановительным потенциалом — НАДН и ФАДН2. На заключительном этапе восстановительные эквиваленты окисляются в электрон-транспортной цепи, локализованной во внутренней мембране митохондрий. Перенос электрона в цепи завершается восстановлением кислорода до воды. В процессе электронного транспорта на мембране образуется электрохимический протонный градиент ΔµἨ, энергия которого используется для синтеза АТФ из АДФ и Фн. Процесс, в котором работа дыхательной цепи сопряжена с синтезом АТФ, получил название окислительного фосфорилирования. Именно в этом процессе синтезируется основная масса АТФ, образуемого при дыхании. И у растений, и у животных дыхание выполняет три основные функции. Во-первых, освобождаемая при окислении углеводов энергия преобразуется в конвертируемые формы клеточной энергии — ΔµἨ и АТФ. Вторая, не менее важная функция — снабжение клетки метаболитами, которые образуются в ходе окисления глюкозы и используются в разнообразных биосинтезах. Третья функция связана с термогенезом, т. е. рассеиванием энергии в виде тепла. Процесс дыхания принципиально сходен у животных и растений, но у последних имеет свои особенности. Все вместе они отражают пластичность растительного метаболизма и связаны с функционированием, наряду с основными, альтернативных ферментов и реакций. Наличие альтернативных путей расширяет адаптивные возможности растений, но усложняет (с точки зрения исследователя) систему регуляции метаболических процессов.
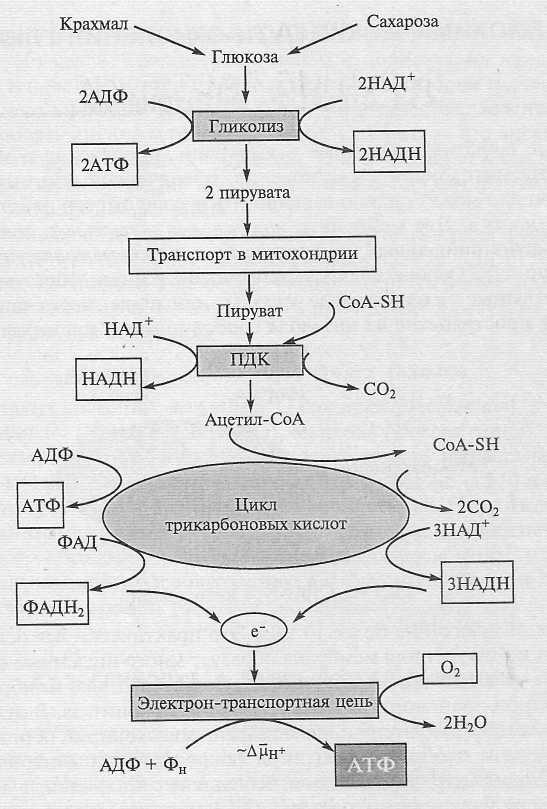
Рис. 1. Основные этапы дыхания
Окисление глюкозы в процессе гликолиза сопровождается восстановлением двух молекул НАД+, синтезом двух молекул АТФ и завершается образованием двух молекул пирувата. В митохондриях пируват подвергается полному окислению до СО2 в реакциях, катализируемых пируватдегидрогеназным комплексом (ПДК) и ферментами цикла трикарбоновых кислот (ЦТК). В этих процессах образуются 4НАДН, 1ФАДН2, а также одна молекула АТФ. Восстановительные эквиваленты окисляются, отдавая электроны в электрон-транспортную цепь, локализованную во внутренней митохондриальной мембране. Электронный транспорт приводит к восстановлению кислорода до воды и сопряжен с синтезом основной массы АТФ в процессе окислительного фосфорилирования.
Основные
события, связанные с дыханием, происходят
в митохондриях. Растительные митохондрии,
как правило, сферической или цилиндрической
формы, их число может сильно варьировать
в зависимости от метаболической
активности клетки. Две мембраны, наружная
и внутренняя, делят
митохондрию на два функциональных
компартмента — межмембранное
пространство и матрикс (рис.
2).
 Рис.
2. Структура митохондрий
Особые
белки, называемые поринами, образуют
в наружной мембране крупные гидрофильные
каналы, или поры, через которые в
межмембранное пространство из
цитозоля свободно могут проникать
соединения с молекулярной массой не
более 10 кДа. Это практически все основные
метаболиты клетки. Внутренняя мембрана
образует многочисленные складки,
кри-сты, которые увеличивают ее
поверхность. Во внутреннюю мембрану
интегрированы электрон-транспортная
цепь (ЭТЦ) и АТФ-синтаза. В отличие от
других клеточных мембран внутренняя
мембрана митохондрий обогащена белком
(75 %) и
содержит особый фосфолипид
(дифосфатидилглицерол) — кардиолипин.
Она пропускает газы, воду и небольшие
липофильные молекулы, но непроницаема
для заряженных молекул и ионов, что
является обязательным условием ее
функционирования как сопрягающей
мембраны. Однако в мембране есть
белки — транспортеры, с помощью которых
возможен обмен метаболитами между
матриксом и цитозолем . Матрикс, т. е.
окруженное внутренней мембраной
пространство, содержит ферменты цикла
трикарбоновых кислот.
Рис.
2. Структура митохондрий
Особые
белки, называемые поринами, образуют
в наружной мембране крупные гидрофильные
каналы, или поры, через которые в
межмембранное пространство из
цитозоля свободно могут проникать
соединения с молекулярной массой не
более 10 кДа. Это практически все основные
метаболиты клетки. Внутренняя мембрана
образует многочисленные складки,
кри-сты, которые увеличивают ее
поверхность. Во внутреннюю мембрану
интегрированы электрон-транспортная
цепь (ЭТЦ) и АТФ-синтаза. В отличие от
других клеточных мембран внутренняя
мембрана митохондрий обогащена белком
(75 %) и
содержит особый фосфолипид
(дифосфатидилглицерол) — кардиолипин.
Она пропускает газы, воду и небольшие
липофильные молекулы, но непроницаема
для заряженных молекул и ионов, что
является обязательным условием ее
функционирования как сопрягающей
мембраны. Однако в мембране есть
белки — транспортеры, с помощью которых
возможен обмен метаболитами между
матриксом и цитозолем . Матрикс, т. е.
окруженное внутренней мембраной
пространство, содержит ферменты цикла
трикарбоновых кислот.
ГЛЮКОЗА — ОСНОВНОЙ СУБСТРАТ ДЫХАНИЯ У РАСТЕНИЙ Основным субстратом дыхания у растений являются глюкоза и ее производные, хотя в особых случаях дыхание могут поддерживать белки и жиры, запасенные в семенах. Глюкоза образуется в клетках растений при гидролизе крахмала и сахарозы — продуктов фотосинтеза. Крахмал представляет собой смесь двух полисахаридов — амилозы и амилопектина. Молекулы амилозы — это длинные, неразветвленные цепи α-D-глюкопиранозных остатков, соединенных гликозидными α(1→4)-связями. Молекулы амилопектина также представлены цепями α -D-глюкопиранозных остатков, которые в точке ветвления образуют а(1→6)-связь. Крахмал как запасный полисахарид накапливается в хлоропластах и пластидах гетеротрофных тканей. Некоторые растения — топинамбур (Heliantus tuberosus), георгин (Dahlia sp.) в качестве запасных углеводов могут использовать инулин и гемицеллюлозы. Сахароза — это дисахарид, образованный остатками глюкозы и фруктозы. Она синтезируется в цитозоле, из фотосинтезирующих клеток по апопласту листа и сосудам флоэмы транспортируется в другие органы растения. Крахмал расщепляется до моносахаридов при участии ряда ферментов (α- и β-амилазы, α-1,6-глюкозидазы, крахмалфосфорилазы и др.) с образованием D-глюкозы или D-глюкозо-1-фосфата. Распад сахарозы может идти при обращении реакций ее синтеза, но в основном происходит в результате гидролиза при участии фермента инвертазы: сахароза + Н2О → фруктоза + глюкоза
В геноме таких растений, как томат (Lycopersicon esculentum), кукуруза (Zea mays), арабидопсис (Arabidopsis thaliana), морковь (Dancus carota), обнаружено целое семейство ядерных генов, кодирующих разные изоформы инвертазы. Например, у моркови кислые инвертазы (оптимум рН 4,5 — 5,0) в пяти разных изоформах присутствуют в вакуоли и клеточной стенке. В цитозоле есть нейтральная инвертаза (оптимум рН 7,0—8,0), которая также может иметь несколько изоформ. Таким образом, у растений гидролиз сахарозы может идти в разных клеточных компартментах и контролируется сложным образом через активность инвертаз, обладающих разными свойствами.
Брожение- процесс анаэробного расщепления органических веществ, преимущественно углеводов, происходящий под влиянием микроорганизмов или выделенных из них ферментов. В ходе брожение в результате сопряженных окислительно-восстановительных реакций освобождается энергия, необходимая для жизнедеятельности микроорганизмов, и образуются химические соединения, которые микроорганизмы используют для биосинтеза аминокислот, белков, органических кислот, жиров и др. компонентов тела. Одновременно накапливаются конечные продукты брожение. В зависимости от их характера различают брожение спиртовое, молочнокислое, маслянокислое, пропионовокислое, ацетоно-бутиловое, ацетоно-этиловое и др. виды. Характер брожение, его интенсивность, количественные соотношения конечных продуктов, а также направление брожение зависят от особенностей его возбудителя и условий, при которых брожение протекает (pH, аэрация, субстрат и др.).
Спиртовое Брожение. В 1836 французский учёный Каньяр де ла Тур установил, что спиртовое брожение связано с ростом и размножением дрожжей. Химическое уравнение спиртового брожение: C6H12O6 ® 2C2H5OH + 2CO2 было дано французскими химиками А. Лавуазье (1789) и Ж. Гей-Люссаком (1815). Л. Пастер пришёл к выводу (1857), что спиртовое брожение могут вызывать только живые дрожжи в анаэробных условиях («брожение — это жизнь без воздуха»). В противовес этому немецкий учёный Ю. Либих упорно настаивал на том, что брожение происходит вне живой клетки. На возможность бесклеточного спиртового брожение впервые (1871) указала русский врач-биохимик М. М. Манассеина.
Немецкий химик Э. Бухнер в 1897, отжав под большим давлением дрожжи, растёртые с кварцевым песком, получил бесклеточный сок, сбраживающий сахар с образованием спирта иCO2. При нагревании до 50°C и выше сок утрачивал бродильные свойства. Всё это указывало на ферментативную природу активного начала, содержащегося в дрожжевом соке. Русский химик Л. А. Иванов обнаружил (1905), что добавленные к дрожжевому соку фосфаты в несколько раз повышают скорость брожение.
Исследования
отечественных биохимиков А. И. Лебедева,
С. П. Костычева, Я. О. Парнаса и немецких
биохимиков К. Нейберга, Г. Эмбдена, О.
Мейергофа и др. подтвердили,
что фосфорная кислота
участвует в важнейших этапах
спиртового брожение
 В дальнейшем многие исследователи
детально изучили ферментативную природу
и механизм спиртового брожение (см.
схему). Первая реакция превращения глюкозы при
спиртовом брожение —
присоединение к глюкозе под
влиянием фермента глюкокиназы
остатка фосфорной кислоты
отаденозинтрифосфорной кислоты
(АТФ, см. Аденозинфосфорные
кислоты).
При этом образуются аденозиндифосфорная кислота
(АДФ) и глюкозо-6-фосфорная кислотата.
Последняя под действием
фермента глюкозофосфати-зомеразы превращается
в фруктозо-6-фосфорную кислоту,
которая, получая от новой молекулы АТФ
(при участии фермента фосфофруктокиназы)
ещё один остаток фосфорной кислоты,
превращается в фруктозо-1,6-дифосфорную
кислоту. (Эта и следующая реакции,
обозначенные встречными стрелками,
обратимы, т. е. их направление зависит
от условий — концентрации фермента, pH
и др.) Под влиянием фермента
кетозо-1-фосфатальдолазы
фруктозо-1,6-дифосфорная кислота
расщепляется на глицеринальдегидфосфорную
и диоксиацетонфосфорную кислоты которые
могут превращаться друг в друга под
действием фермента триозофосфатизомеразы.
Глицеринальдегидфосфорная кислота,
присоединяя молекулу
неорганической фосфорной кислоты
и окисляясь под действием
фермента дегидрогеназы фосфоглицеринальдегида,
активной группой которого у дрожжей
является никотинамидадениндинуклеотид
(НАД), превращается в 1,3-дифосфоглицериновую
кислоту. Молекула диоксиацетонфосфорной
кислоты под действием триозофосфатизомеразы
даёт вторую молекулу глицеринальдегидфосфорной
кислоты, также подвергающуюся окислению
до 1,3-дифосфоглицериновой кислоты;
последняя, отдавая АДФ (под действием
фермента фосфоглицераткиназы) один
остаток фосфорной кислоты,
превращается в З-фосфоглицериновую
кислоту, которая под действием фермента
фосфоглицеро-мутазы превращается в
2-фосфоглицериновую кислоту, а она под
влиянием фермента фосфопируват-гидратазы
— в фосфоенол-пировиноградную кислоту.
Последняя при участии фермента
пируваткиназы передаёт остаток фосфорнойкислоты
молекуле АДФ, в результате чего образуется
молекула АТФ и молекула енолпировиноградной
кислоты, которая весьма нестойка и
переходит в пировиноградную кислоту.
Эта кислота при участии имеющегося в
дрожжах фермента пируватдекарбоксилазы
расщепляется на уксусный альдегид и
двуокись углерода. Уксусный
альдегид, реагируя с образовавшейся
при окислении глицеринальдегидфосфорной
кислоты восстановленной формой
никотинамидадениндинуклеотида (НАД-Н),
при участии фермента алкогольдегидрогеназы
превращается в этиловый спирт. Суммарно
уравнение спиртового брожение может
быть представлено в следующем
виде:
C6H12O6 +
2H3PO4 +
2АДФ ® 2CH3CH2OH
+ 2CO2 +
2АТФ.
Т. о., при сбраживании 1 моля глюкозы образуются
2 моля этилового
спирта, 2 моля CO2,
а также в результате фосфорилирования 2 молей АДФ
образуются 2 моля АТФ.
Термодинамические расчёты показывают,
что при спиртовом брожение превращение
1 моля глюкозы может
сопровождаться уменьшением свободной
энергии примерно на 210 кдж (50
000 кал),
т. е. энергия,
аккумулированная в 1 моле этилового
спирта, на 210 кдж (50
000 кал)
меньше энергии 1 моля глюкозы. При
образовании 1 моля АТФ
(макроэргических — богатых энергией
фосфатных соединений) используется
42 кдж (10
000 кал).
Следовательно, значительная часть
энергии, освобождающейся при спиртовом
брожение,
запасается в виде АТФ, обеспечивающей
разнообразные энергетические потребности
дрожжевых клеток. Такое же биологическое
значение имеет процесс брожение и
у др. микроорганизмов. При полном сгорании
1 моля глюкозы (с
образованием CO2 и
H2O)
изменение свободной энергии достигает
2,87 Мдж (686
000 кал).
Иначе говоря, дрожжевая клетка использует
лишь 7% энергии глюкозы. Это
показывает малую эффективность анаэробных
процессов по сравнению с процессами,
идущими в присутствии кислорода. При
наличии кислорода спиртовое брожение угнетается
или прекращается и дрожжи получают
энергию для жизнедеятельности в процессе
дыхания. Наблюдается тесная связь
между брожением и
дыханием микроорганизмов, растений и
животных. Ферменты, участвующие в
спиртовом брожение,
имеются также в тканях животных и
растений. Во многих случаях первые этапы
расщепления сахаров, вплоть
до образования пировиноградной кислоты,
— общие для брожение и
дыхания. Большее значение процесс
анаэробного распада глюкозы имеет
и при сокращении мышц , первые этапы
этого процесса также сходны с начальными
реакциями спиртового брожение.
Сбраживание углеводов (глюкозы, ферментативных
гидролизатов крахмала, кислотных
гидролизатов древесины) используется
во многих отраслях промышленности: для
получения этилового спирта, глицерина
и др. технических и пищевых продуктов.
На спиртовом Брожение
основаны
приготовление теста в хлебопекарной
промышленности, виноделие и
пивоварение.
Молочнокислое Брожение. Молочнокислые
бактерии подразделяют на 2 группы —
гомоферментативные и гетероферментативные.
Гомоферментативные бактерии (например,
Lactobacillus delbrückii) расщепляют моносахариды
с образованием двух молекул молочной
кислоты в соответствии с суммарным
уравнением:
C6H12O6 =
2CH3CHOH·COOH.
Гетероферментативные бактерии (например,
Bacterium lactis aerogenes) ведут сбраживание с
образованием молочной кислоты, уксусной
кислоты, этилового спирта и CO2,
а также образуют небольшое количество
ароматических. веществ — диацетила,
эфиров и т.д.
При молочнокислом брожение превращение
углеводов, особенно на первых этапах,
близко к реакциям спиртового брожение,
за исключением декарбоксилирования
пировиноградной кислоты, которая
восстанавливается до молочной кислоты
за счёт водорода, получаемого от НАД-Н.
Гомоферментативное
молочнокислое брожение используется
для получения молочной кислоты, при
изготовлении различных кислых молочных
продуктов, хлеба и в силосовании кормов
в сельском хозяйстве. Гетероферментативное
молочнокислое брожение происходит
при консервировании различных плодов
и овощей путём
квашения.
Маслянокислое Брожение. Сбраживание
углеводов с преимущественным образованием
масляной кислоты производят многие
анаэробные бактерии, относящиеся к роду
Clostridium. Первые этапы расщепления углеводов
при маслянокислом Брожение аналогичны
соответстветственным этапам
спиртового брожение,
вплоть до образования пировиноградной
кислоты, из которой при
маслянокислом брожение образуется
ацетил-кофермент A (CH3CO-KoA).
Ацетил-KoA может служить предшественником
масляной кислоты, подвергаясь следующим
превращениям:
В дальнейшем многие исследователи
детально изучили ферментативную природу
и механизм спиртового брожение (см.
схему). Первая реакция превращения глюкозы при
спиртовом брожение —
присоединение к глюкозе под
влиянием фермента глюкокиназы
остатка фосфорной кислоты
отаденозинтрифосфорной кислоты
(АТФ, см. Аденозинфосфорные
кислоты).
При этом образуются аденозиндифосфорная кислота
(АДФ) и глюкозо-6-фосфорная кислотата.
Последняя под действием
фермента глюкозофосфати-зомеразы превращается
в фруктозо-6-фосфорную кислоту,
которая, получая от новой молекулы АТФ
(при участии фермента фосфофруктокиназы)
ещё один остаток фосфорной кислоты,
превращается в фруктозо-1,6-дифосфорную
кислоту. (Эта и следующая реакции,
обозначенные встречными стрелками,
обратимы, т. е. их направление зависит
от условий — концентрации фермента, pH
и др.) Под влиянием фермента
кетозо-1-фосфатальдолазы
фруктозо-1,6-дифосфорная кислота
расщепляется на глицеринальдегидфосфорную
и диоксиацетонфосфорную кислоты которые
могут превращаться друг в друга под
действием фермента триозофосфатизомеразы.
Глицеринальдегидфосфорная кислота,
присоединяя молекулу
неорганической фосфорной кислоты
и окисляясь под действием
фермента дегидрогеназы фосфоглицеринальдегида,
активной группой которого у дрожжей
является никотинамидадениндинуклеотид
(НАД), превращается в 1,3-дифосфоглицериновую
кислоту. Молекула диоксиацетонфосфорной
кислоты под действием триозофосфатизомеразы
даёт вторую молекулу глицеринальдегидфосфорной
кислоты, также подвергающуюся окислению
до 1,3-дифосфоглицериновой кислоты;
последняя, отдавая АДФ (под действием
фермента фосфоглицераткиназы) один
остаток фосфорной кислоты,
превращается в З-фосфоглицериновую
кислоту, которая под действием фермента
фосфоглицеро-мутазы превращается в
2-фосфоглицериновую кислоту, а она под
влиянием фермента фосфопируват-гидратазы
— в фосфоенол-пировиноградную кислоту.
Последняя при участии фермента
пируваткиназы передаёт остаток фосфорнойкислоты
молекуле АДФ, в результате чего образуется
молекула АТФ и молекула енолпировиноградной
кислоты, которая весьма нестойка и
переходит в пировиноградную кислоту.
Эта кислота при участии имеющегося в
дрожжах фермента пируватдекарбоксилазы
расщепляется на уксусный альдегид и
двуокись углерода. Уксусный
альдегид, реагируя с образовавшейся
при окислении глицеринальдегидфосфорной
кислоты восстановленной формой
никотинамидадениндинуклеотида (НАД-Н),
при участии фермента алкогольдегидрогеназы
превращается в этиловый спирт. Суммарно
уравнение спиртового брожение может
быть представлено в следующем
виде:
C6H12O6 +
2H3PO4 +
2АДФ ® 2CH3CH2OH
+ 2CO2 +
2АТФ.
Т. о., при сбраживании 1 моля глюкозы образуются
2 моля этилового
спирта, 2 моля CO2,
а также в результате фосфорилирования 2 молей АДФ
образуются 2 моля АТФ.
Термодинамические расчёты показывают,
что при спиртовом брожение превращение
1 моля глюкозы может
сопровождаться уменьшением свободной
энергии примерно на 210 кдж (50
000 кал),
т. е. энергия,
аккумулированная в 1 моле этилового
спирта, на 210 кдж (50
000 кал)
меньше энергии 1 моля глюкозы. При
образовании 1 моля АТФ
(макроэргических — богатых энергией
фосфатных соединений) используется
42 кдж (10
000 кал).
Следовательно, значительная часть
энергии, освобождающейся при спиртовом
брожение,
запасается в виде АТФ, обеспечивающей
разнообразные энергетические потребности
дрожжевых клеток. Такое же биологическое
значение имеет процесс брожение и
у др. микроорганизмов. При полном сгорании
1 моля глюкозы (с
образованием CO2 и
H2O)
изменение свободной энергии достигает
2,87 Мдж (686
000 кал).
Иначе говоря, дрожжевая клетка использует
лишь 7% энергии глюкозы. Это
показывает малую эффективность анаэробных
процессов по сравнению с процессами,
идущими в присутствии кислорода. При
наличии кислорода спиртовое брожение угнетается
или прекращается и дрожжи получают
энергию для жизнедеятельности в процессе
дыхания. Наблюдается тесная связь
между брожением и
дыханием микроорганизмов, растений и
животных. Ферменты, участвующие в
спиртовом брожение,
имеются также в тканях животных и
растений. Во многих случаях первые этапы
расщепления сахаров, вплоть
до образования пировиноградной кислоты,
— общие для брожение и
дыхания. Большее значение процесс
анаэробного распада глюкозы имеет
и при сокращении мышц , первые этапы
этого процесса также сходны с начальными
реакциями спиртового брожение.
Сбраживание углеводов (глюкозы, ферментативных
гидролизатов крахмала, кислотных
гидролизатов древесины) используется
во многих отраслях промышленности: для
получения этилового спирта, глицерина
и др. технических и пищевых продуктов.
На спиртовом Брожение
основаны
приготовление теста в хлебопекарной
промышленности, виноделие и
пивоварение.
Молочнокислое Брожение. Молочнокислые
бактерии подразделяют на 2 группы —
гомоферментативные и гетероферментативные.
Гомоферментативные бактерии (например,
Lactobacillus delbrückii) расщепляют моносахариды
с образованием двух молекул молочной
кислоты в соответствии с суммарным
уравнением:
C6H12O6 =
2CH3CHOH·COOH.
Гетероферментативные бактерии (например,
Bacterium lactis aerogenes) ведут сбраживание с
образованием молочной кислоты, уксусной
кислоты, этилового спирта и CO2,
а также образуют небольшое количество
ароматических. веществ — диацетила,
эфиров и т.д.
При молочнокислом брожение превращение
углеводов, особенно на первых этапах,
близко к реакциям спиртового брожение,
за исключением декарбоксилирования
пировиноградной кислоты, которая
восстанавливается до молочной кислоты
за счёт водорода, получаемого от НАД-Н.
Гомоферментативное
молочнокислое брожение используется
для получения молочной кислоты, при
изготовлении различных кислых молочных
продуктов, хлеба и в силосовании кормов
в сельском хозяйстве. Гетероферментативное
молочнокислое брожение происходит
при консервировании различных плодов
и овощей путём
квашения.
Маслянокислое Брожение. Сбраживание
углеводов с преимущественным образованием
масляной кислоты производят многие
анаэробные бактерии, относящиеся к роду
Clostridium. Первые этапы расщепления углеводов
при маслянокислом Брожение аналогичны
соответстветственным этапам
спиртового брожение,
вплоть до образования пировиноградной
кислоты, из которой при
маслянокислом брожение образуется
ацетил-кофермент A (CH3CO-KoA).
Ацетил-KoA может служить предшественником
масляной кислоты, подвергаясь следующим
превращениям:
 Маслянокислое Брожение применялось
для получения масляной кислоты из
крахмала.
Ацетоно-бутиловое брожение бактерии
Clostridium acetobutylicum сбраживают углеводы с
преим. образованием бутилового спирта
(CH3CH2CH2CH2OH)
и ацетона (CH3COCH3).
При этом образуются также в сравнительно
небольших количествах водород, CO2,
уксусная, масляная кислоты, этиловый
спирт. Первые этапы расщепления углеводов
те же, что и при спиртовом брожении. Бутиловый
спирт образуется путём восстановления
масляной кислоты:
CH3CH2CH2COOH
+ 4H = CH3CH2CH2CH2OH
+ H2O.
Ацетон же образуется декарбоксилированием
ацетоуксусной кислоты, которая получается
в результате конденсации двух молекул
уксусной кислоты. Исследованиями В. Н.
Шапошникова показано, что
ацетоно-бутиловое брожение (как
и ряд др., например пропионовокислое,
маслянокислое) в опытах с растущей
культурой происходит в две фазы. В первую
фазу брожение параллельно
с нарастанием биомассы накапливаются
уксусная и масляная кислоты; во вторую
фазу образуются преимущественно ацетон
и бутиловый спирт. При ацетоно-бутиловом
брожение сбраживаются
моносахариды, дисахариды и полисахариды
— крахмал, инсулин, но не сбраживаются
клетчатка и гемицеллюлоза.
Ацетоно-бутиловое брожение использовалось
для промышленного получения бутилового
спирта и ацетона, применяемых в химической
и лакокрасочной
промышленности.
Сбраживание белков. Некоторые
бактерии из рода Clostridium —
гнилостные анаэробы — способны
сбраживать не только углеводы, но и
аминокислоты. Эти бактерии более
приспособлены к использованию белков, расщепляемых
ими при помощи протеолитических ферментов
до аминокислот, которые затем
подвергаются брожение. Процесс
сбраживания белков имеет
значение в круговороте веществ в
природе.
Пропионовокислое Брожение. Основные
продукты пропионовокислого брожение,
вызываемого несколькими видами бактерий
из рода Propionibacterium, — пропионовая
(CH3CH2OH)
и уксусная кислоты и CO2.
Химизм пропионовокислого брожение сильно
изменяется в зависимости от условий.
Это, по-видимому, объясняется способностью
пропионовых бактерий перестраивать
обмен веществ, например в зависимости
от аэрации. При доступе кислорода они
ведут окислительный процесс, а в его
отсутствии расщепляют гексозы
путём брожение. Пропионовые
бактерии способны фиксировать CO2,
при этом из пировиноградной к-ты
и CO2 образуется
щавелевоуксусная к-та, превращающаяся
в янтарную к-ту, из которой декарбоксилированием
образуется пропионовая к-та:
Маслянокислое Брожение применялось
для получения масляной кислоты из
крахмала.
Ацетоно-бутиловое брожение бактерии
Clostridium acetobutylicum сбраживают углеводы с
преим. образованием бутилового спирта
(CH3CH2CH2CH2OH)
и ацетона (CH3COCH3).
При этом образуются также в сравнительно
небольших количествах водород, CO2,
уксусная, масляная кислоты, этиловый
спирт. Первые этапы расщепления углеводов
те же, что и при спиртовом брожении. Бутиловый
спирт образуется путём восстановления
масляной кислоты:
CH3CH2CH2COOH
+ 4H = CH3CH2CH2CH2OH
+ H2O.
Ацетон же образуется декарбоксилированием
ацетоуксусной кислоты, которая получается
в результате конденсации двух молекул
уксусной кислоты. Исследованиями В. Н.
Шапошникова показано, что
ацетоно-бутиловое брожение (как
и ряд др., например пропионовокислое,
маслянокислое) в опытах с растущей
культурой происходит в две фазы. В первую
фазу брожение параллельно
с нарастанием биомассы накапливаются
уксусная и масляная кислоты; во вторую
фазу образуются преимущественно ацетон
и бутиловый спирт. При ацетоно-бутиловом
брожение сбраживаются
моносахариды, дисахариды и полисахариды
— крахмал, инсулин, но не сбраживаются
клетчатка и гемицеллюлоза.
Ацетоно-бутиловое брожение использовалось
для промышленного получения бутилового
спирта и ацетона, применяемых в химической
и лакокрасочной
промышленности.
Сбраживание белков. Некоторые
бактерии из рода Clostridium —
гнилостные анаэробы — способны
сбраживать не только углеводы, но и
аминокислоты. Эти бактерии более
приспособлены к использованию белков, расщепляемых
ими при помощи протеолитических ферментов
до аминокислот, которые затем
подвергаются брожение. Процесс
сбраживания белков имеет
значение в круговороте веществ в
природе.
Пропионовокислое Брожение. Основные
продукты пропионовокислого брожение,
вызываемого несколькими видами бактерий
из рода Propionibacterium, — пропионовая
(CH3CH2OH)
и уксусная кислоты и CO2.
Химизм пропионовокислого брожение сильно
изменяется в зависимости от условий.
Это, по-видимому, объясняется способностью
пропионовых бактерий перестраивать
обмен веществ, например в зависимости
от аэрации. При доступе кислорода они
ведут окислительный процесс, а в его
отсутствии расщепляют гексозы
путём брожение. Пропионовые
бактерии способны фиксировать CO2,
при этом из пировиноградной к-ты
и CO2 образуется
щавелевоуксусная к-та, превращающаяся
в янтарную к-ту, из которой декарбоксилированием
образуется пропионовая к-та:
 Существуют брожение,
которые сопровождаются и восстановительными
процессами. Примером такого
«окислительного» брожение служит
лимоннокислое брожение. Многие
плесневые грибы сбраживают сахара с
образованием лимонной кислоты. Наиболее
активные штаммы Aspergillus niger превращают
до 90% потребленного сахара в
лимонную кислоту. Значительная часть
лимонной кислоты, используемой в пищевой
промышленности, производится
микробиологическим путём — глубинным
и поверхностным культивированием
плесневых грибов.
Иногда по традиции и чисто окислительные
процессы, осуществляемые микроорганизмами,
называется брожение. Примерами
таких процессов могут служить уксуснокислое
и глюконовокислое брожение.
Уксуснокислое Брожение. Бактерии,
относящиеся к роду Acetobacter, окисляют
этиловый спирт в уксусную кислоту в
соответствии с суммарной реакцией:
Существуют брожение,
которые сопровождаются и восстановительными
процессами. Примером такого
«окислительного» брожение служит
лимоннокислое брожение. Многие
плесневые грибы сбраживают сахара с
образованием лимонной кислоты. Наиболее
активные штаммы Aspergillus niger превращают
до 90% потребленного сахара в
лимонную кислоту. Значительная часть
лимонной кислоты, используемой в пищевой
промышленности, производится
микробиологическим путём — глубинным
и поверхностным культивированием
плесневых грибов.
Иногда по традиции и чисто окислительные
процессы, осуществляемые микроорганизмами,
называется брожение. Примерами
таких процессов могут служить уксуснокислое
и глюконовокислое брожение.
Уксуснокислое Брожение. Бактерии,
относящиеся к роду Acetobacter, окисляют
этиловый спирт в уксусную кислоту в
соответствии с суммарной реакцией:
![]() Промежуточное соединение при окислении
спирта в уксусную кислоту — уксусный
альдегид. Многие уксуснокислые бактерии,
кроме окисления спирта в уксусную
кислоту, осуществляют окисление глюкозы в
глюконовую и кетоглюконовую
кислоты.
Глюконовокислое Брожение осуществляют
и некоторые плесневые грибы, способные
окислять альдегидную группу глюкозы, превращая
последнюю в глюконовую
кислоту:
Промежуточное соединение при окислении
спирта в уксусную кислоту — уксусный
альдегид. Многие уксуснокислые бактерии,
кроме окисления спирта в уксусную
кислоту, осуществляют окисление глюкозы в
глюконовую и кетоглюконовую
кислоты.
Глюконовокислое Брожение осуществляют
и некоторые плесневые грибы, способные
окислять альдегидную группу глюкозы, превращая
последнюю в глюконовую
кислоту:
![]() Кальциевая соль
глюконовой кислоты служит хорошим
источником кальция для людей и животных.
Кальциевая соль
глюконовой кислоты служит хорошим
источником кальция для людей и животных.