

14-15 Матричные биосинтезы
.pdfМАТРИЧНЫЕ БИОСИНТЕЗЫ
Основной фигурой матричных биосинтезов являются нуклеиновые кислоты. Они
представляют собой полимерные молекулы, в состав которых входят азотистые ос-
нования пяти типов, пентозы двух типов и остатки фосфорной кислоты.
Азотистые основания в нуклеиновых кислотах могут быть пуриновыми (аденин,
гуанин) и пиримидиновыми (цитозин, урацил, тимин).
Взависимости от строения углевода выделяют рибонуклеиновые кислоты – содержат рибозу (РНК), и дезоксирибонуклеиновые кислоты – содержат дезоксирибо-
зу (ДНК).
ОСНОВНОЙ ПОСТУЛАТ МОЛЕКУЛЯРНОЙ БИОЛОГИИ
Вподавляющем большинстве случаев передача наследственной информации
от материнской клетки к дочерней осуществляется при помощи ДНК. Для использо-
вания генетической информации самой клеткой необходимы РНК, образуемые на матрице ДНК. Далее РНК непосредственно участвуют на всех этапах синтеза белко-
вых молекул, обеспечивающих структуру и деятельность клетки.
На вышесказанном основана центральная догма молекулярной биологии, со-
гласно которой перенос генети-
ческой информации осуществля-
ется только от нуклеиновой кислоты (ДНК и РНК). Получателем
информации может быть другая
нуклеиновая кислота (ДНК или РНК) и белок.
СТРОЕНИЕ ДЕЗОКСИРИБОНУКЛЕИНОВОЙ КИСЛОТЫ
ДНК – наиболее важная часть
хромосом: две двухцепочечные
молекулы ДНК образуют одну хромосому. Наиболее хорошо
хромосомы видны перед митозом
и во время его. В покоящихся клетках хромосомный материал
выглядит нечетко и распределен
по всему ядру. В таком состоя-
нии он получил название "хрома-
тин". В составе хроматина выде-
ляют 60% белка (гистоны и кислые белки), 35% ДНК и около 5% РНК.
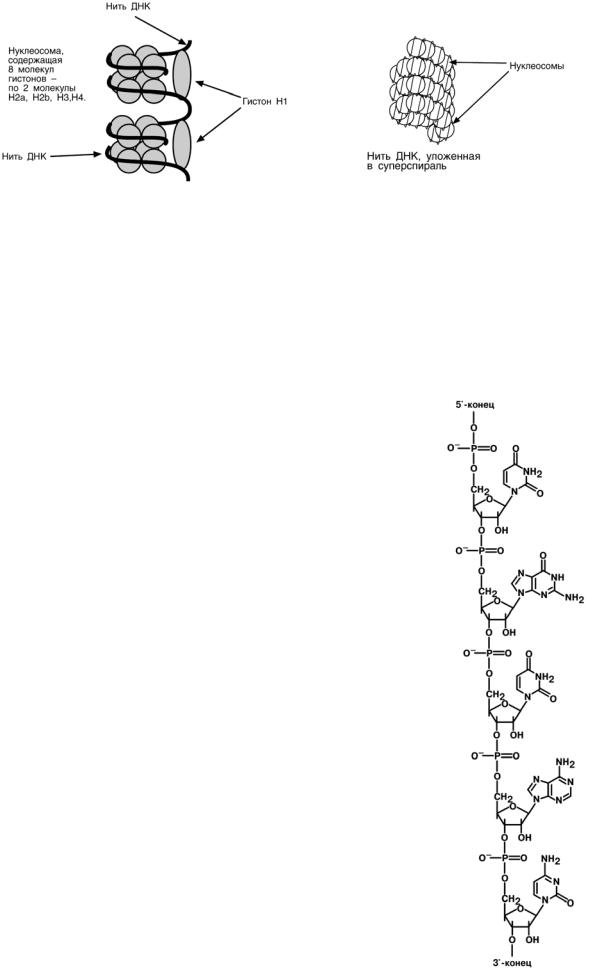
2
Хроматин уложен в виде сферических частиц – нуклеосом, соединенных друг с
другом нитью ДНК. Нуклеосома представляет собой комплекс участка молекулы ДНК и восьми молекул гистонов. В составе нуклеосомы находятся по 2 молекулы гисто-
нов Н2α, Н2β, Н3, Н4. Нить ДНК последовательно контактируя с гистонами Н2α, Н2β,
Н4, Н3, Н3, Н4, Н2β, Н2α, наматывается на гистоновое ядро, которое "маскирует" 146 пар оснований ДНК. Гистон Н1 связывается с нуклеосомой на участке входа и
выхода ДНК, "склеивая" 2 оборота и "маскируя" еще 20 пар оснований. Всего за-
маскировано 166 пар оснований. Кроме нуклеосом, в ядре присутствуют еще 2
структуры: фибриллы диаметром 10 нм, состоящие из цепочки нуклеосом, и волокна, диаметром 30 нм, образующиеся при закручивании фибрилл в спираль. На виток спирали приходится 6-7 нуклеосом. Участок ДНК между нуклеосомами называется спейсерным (англ: space – пространство), его длина варьирует в зависимости от вида организма и типа клеток. У человека она составляет около 50 пар нуклеотидов.
Благодаря наличию нуклеосом достигается уменьшение размеров хромосомы в 7 раз, далее происходит укладка в суперспираль и „суперсуперспираль". Таким образом, благодаря гистонам размеры ДНК уменьшаются в тысячи раз: если длина ДНК достигает 6-9 см (10-1), то размеры хромосом – всего несколько микрометров (10-6).
Хроматин может быть активным (эухроматин) и неактивным (гетерохроматин). Активный хроматин содержит активные гены, т.е. те гены, с которых считывается информация. В активном хроматине нуклеосомная структура изменена или вообще отсутству-
ет, благодаря чему ДНК становится доступной для со-
ответствующих ферментов.
СТРОЕНИЕ РИБОНУКЛЕИНОВЫХ
КИСЛОТ
Рибонуклеиновая кислота (РНК) представляет со-
бой последовательность рибонуклеозидмонофосфа-
тов, связанных друг с другом 5’-3’-фосфодиэфирными связями. РНК отличается от ДНК однонитевой струк-
3
турой, наличием урацила вместо тимина и рибозы вместо дезоксирибозы
В клетке присутствует четыре типа РНК:
Рибосомальные РНК (рРНК) у прокариот и эукариот различны и отличаются величиной седиментации (S, величиной скорости оседания молекулы). У прокариот три
разновидности рРНК: 5S, 16S и 23S. У эукариот четыре разновидности: 5S, 5,8S, 18S
и 28S. Рибосомальные РНК участвуют в построении рибосом, внутриклеточных белоксинтезирующих органелл.
Рибосомы состоят из двух неравных субчастиц, малой и большой.
Упрокариот
•малую (30S) субчастицу образуют белки, 23S-рРНК и 5S-рРНК;
•большую (50S) – белки и 16S-рРНК.
Уэукариот
•малую (40S) субчастицу образуют белки и 18S-рРНК,
•большую (60S) – белки и 5S-, 5,8S-, 28S-рРНК.
Матричные РНК (мРНК) представляют собой линейную последовательность нуклеотидов. К 5’-концу молекулы присоеди-
нен метилгунозиндифосфат, на 3’-конце име-
ется полиадениловая последовательность. Их функция – информационная, т.е. перенос информации о структуре белков от ДНК к месту
их синтеза.
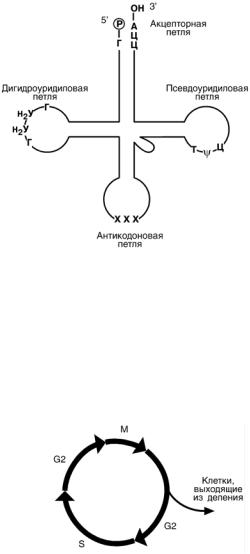
Транспортные РНК (тРНК) бактерий и эу-
кариот включают 73-93 нуклеотида. Они пере-
носят аминокислоты из цитозоля к рибосомам. Вторичная структура тРНК напоминает кле-
верный лист, а третичная – латинскую букву L.
В «клеверном листе» выделяют четыре участка (или ветви, петли), каждый из которых име-
ет собственную функцию: антикодоновый,
псевдоуридиловый, дигидроуридиловый, акцепторный. На 5’-конце тРНК находится гуани-
ловый нуклеотид, на 3’-конце – триплет Ц-Ц-А.
• • Малые РНК – используются для созревания мРНК и некоторых других клеточ-
ных процессов.
РЕПЛИКАЦИЯ ДНК
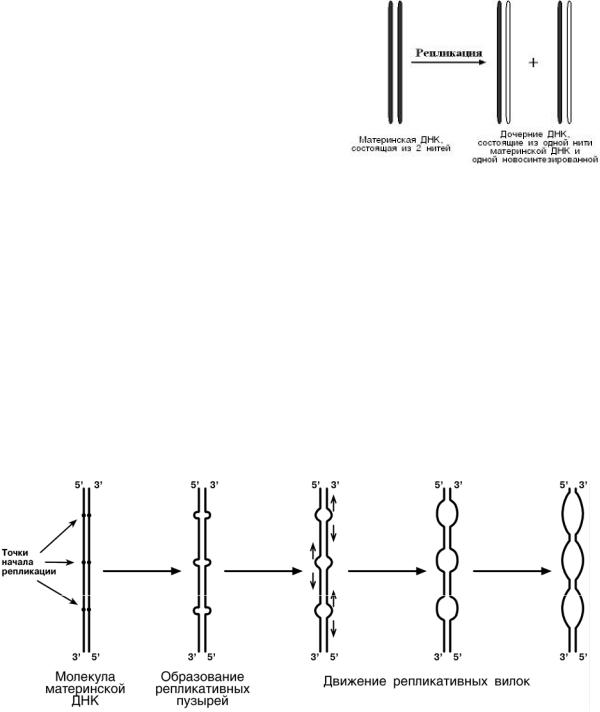
Синтез ДНК в клетке происходит не беспорядоч-
но, а в строго определенный период жизни клетки.
Всего выделяют 4 фазы: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap - интервал),
постсинтетическую (G2).
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой
35-90 кДа, уровень которых меняется в ходе клеточ-
ного цикла. По функции циклины – это активаторные
субъединицы ферментов циклин-зависимых киназ (ЦЗК). Активные комплексы циклин-ЦЗК фосфорили-
руют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
4
Синтез (репликация, удвоение) ДНК про-
исходит в S-фазу клеточного цикла.
Механизм репликации, как установили эксперименты Мэтью Мезельсон и Франклин
Сталь в 1957 г, полуконсервативный, т.е.
на каждой нити материнской ДНК синтезируется дочерняя копия.
Весь процесс репликации идет в S-фазу
клеточного цикла, в то время, когда клетка готовится к делению.
Как матричный биосинтез, репликация
требует наличия нескольких условий:
•Матрица – в ее роли выступает материнская ДНК;
•Растущая цепь – дочерняя ДНК;
•Субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ;
•Источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ;
•Ферменты.
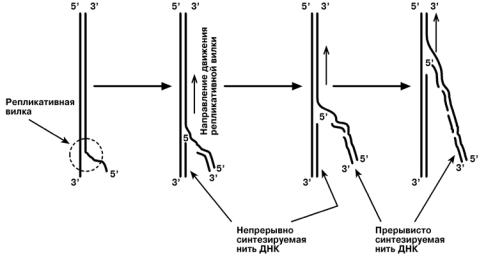
Синтез ДНК начинается в определенных участках, получивших название точка o r i (англ. origin - начало). На каждой ДНК млекопитающих точек o r i насчитывается
около 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием "репликативных пузырей". В каждом таком "пузыре" имеются
две "репликативные вилки", в которых происходит расплетание, раскручивание и
непосредственный синтез ДНК. Репликативные вилки удаляются друг от друга. В целом вся репликация ДНК у эукариот заканчивается за 9 часов.
В каждой репликативной вилке идет синтез ДНК в направлении от 5'-конца к
3'-концу, т.е. 5'-конец новой ДНК остается свободным, следующие нуклеотиды при-
соединяются к 3'-гидроксильной группе предыдущего нуклеотида. Поскольку нити ДНК антипараллельны, то непрерывно синтезируется только одна нить, а именно та, на которой направление движения репликативной вилки совпадает с направлением
3' → 5'.
5
По мере расплетания и движения репликативной вилки на нити открываются
участки, где возможен синтез новой нити в направлении 5' → 3'.
Направление 5' → 3' другой материнской нити ДНК совпадает с направлением движения репликативной вилки. Поэтому синтез дочерней нити (в направлении 5' → 3') возможен только после расплетания части ДНК и освобождения участка для синтеза.
Таким образом, синтез дочерней ДНК на одной из нитей материнской ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
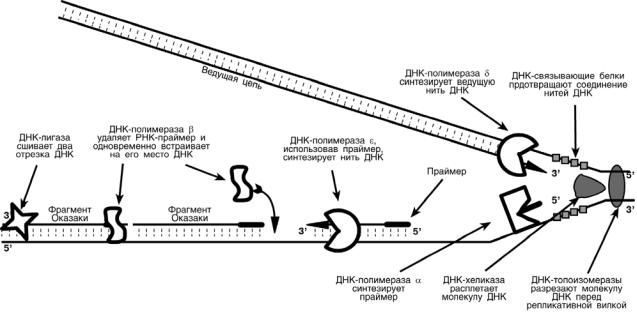
6
В целом для синтеза ДНК необходим ряд ферментов.
Ферменты репликации эукариот и их функция
|
ВИД АКТИВНОСТИ |
ФУНКЦИЯ |
|
|
|
|
|
Топоизомеразы |
Эндонуклеазная |
Разрезание молекулы ДНК для об- |
|
легчения ее расплетания и раскру- |
|||
|
|
чивания |
|
Хеликазы |
Эндонуклеазная |
Раскручивание молекулы ДНК |
|
|
|
|
|
ДНК-связывающие |
|
Стабилизация расплетенных нитей |
|
белки |
|
ДНК |
|
ДНК-полимераза α |
5'-3'–Полимеразная |
Синтез РНК-затравки на основе мо- |
|
лекулы ДНК |
|||
|
|
||
ДНК-полимераза β |
5'-3'–Полимеразная |
Репарация повреждений. |
|
3'-5'–Экзонуклеазная |
|||
|
5'-3'–Экзонуклеазная |
|
|
|
5'-3'–Полимеразная |
Элонгация отстающей цепи дочер- |
|
ДНК-полимераза ε |
ней ДНК на матрице материнской |
||
|
3'-5'–Экзонуклеазная |
ДНК |
|
|
|
||
|
5'-3'–Полимеразная |
Элонгация ведущей цепи дочерней |
|
ДНК-полимераза δ |
3'-5'–Экзонуклеазная |
||
ДНК на матрице материнской ДНК |
|||
|
Экзонуклеазная |
||
|
|
||
ДНК-лигаза |
|
Сшивка фрагментов Оказаки |
|
|
|
|
Роль ферментов репликации ДНК
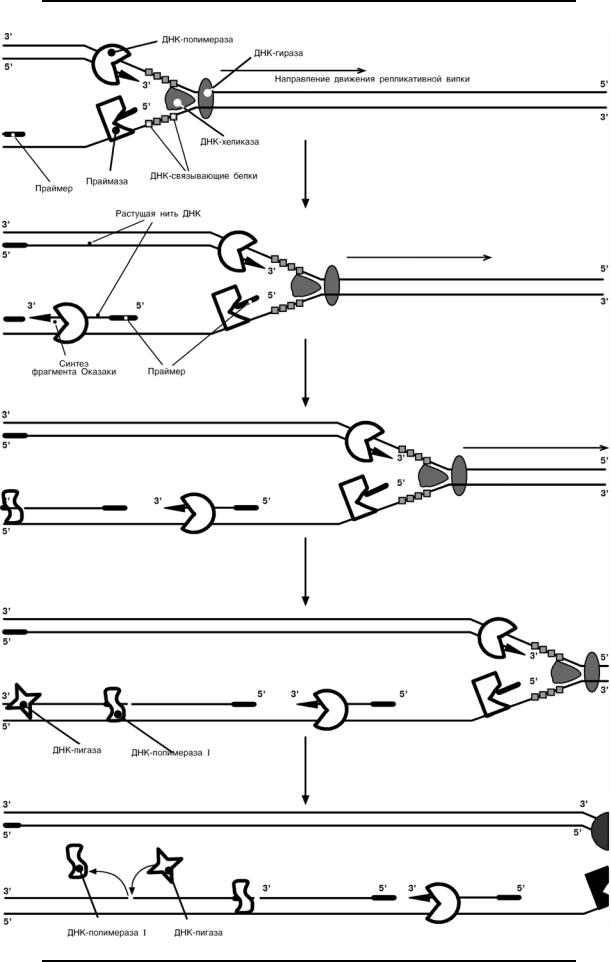
7
Дополнение – движение репликативной вилки и синтез нитей ДНК:

8
ПОВРЕЖДЕНИЯ И РЕПАРАЦИЯ ДНК.
Так как на геном любой неделящейся клетки постоянно оказывает влияние окру-
жающая среда, то вполне вероятны повреждения в составе генома: изменение нуклеотида (например, дезаминирование), сшивки азотистых оснований друг с другом,
разрывы цепей, отрыв пуриновых нуклеотидов и т.п. Такие изменения быстро опре-
деляются специальными ферментами, пораженный участок удаляется экзонуклеа-
зами, заполняется ДНК-полимеразой β и сшивается ДНК-лигазой.
В делящейся клетке мутации могут также возникать во время синтеза ДНК. По-
этому в клетках существует двойная система проверки точности репликации: одна непосредственно при ДНК-полимеразной реакции, другая – анализ уже синтезиро-
ванной ДНК.
ГИБРИДИЗАЦИЯ ДНК– ДНК И ДНК– РНК
Если нагреть раствор ДНК выше температуры 90°С или сдвинуть рН в резко ще-
лочную или резко кислую стороны, то водородные связи между нитями ДНК разру-
шаются, двойная спираль расплетается. Происходит денатурация ДНК или, подругому, плавление. Если удалить агрессивный фактор, то происходит ренатура-
ция или отжиг. При отжиге нити ДНК "отыскивают" комплементарные участки друг у друга и, в конце концов, вновь сворачиваются в двойную спираль.
Если в одной "пробирке" провести плавление и отжиг смеси ДНК человека и мы-
ши, то некоторые участки цепей ДНК мыши будет воссоединяться с комплементар-
ными участками цепей ДНК человека с образованием гибридов. Число таких участков зависит от степени родства видов. Чем ближе виды между собой, тем больше
участков комплементарности нитей ДНК. Это называется гибридизация ДНК-ДНК.
Если в растворе присутствует РНК, то можно осуществить гибридизацию ДНКРНК. Это помогает установить близость определенных последовательностей ДНК с
какой-либо РНК.
Гибридизация ДНК-ДНК и ДНК-РНК используется как эффективное средство в молекулярной генетике.
Например, на основе знания белковой последовательности можно искусственно синтезировать РНК. При гибридизации такой РНК с
образцами ДНК вполне реально определить участок ДНК, ответ-
ственный за синтез исходного белка.
ТРАНСКРИПЦИЯ
Транскрипция (англ. transcription – переписывание), – это биосинтез РНК на мат-
рице ДНК. Биосинтез РНК происходит в участке ДНК, который называется транс-
криптом, с одного края он ограничен промотором (начало), с другого – терминатором (конец).
Как в любом матричном биосинтезе в транскрипции выделяют 5 необходимых элементов:
•матрица – одна из цепей ДНК
•растущая цепь – РНК
•субстрат для синтеза – рибонуклеотиды (УТФ, ГТФ, ЦТФ, АТФ)
•источник энергии – УТФ, ГТФ, ЦТФ, АТФ
•ферменты – РНК-полимеразы.
9
Существует три основных типа РНК-полимераз: для синтеза пре-рРНК (РНК-
полимераза I), для синтеза пре-мРНК (РНК-полимераза II), для синтеза пре-тРНК и
5S-рРНК (РНК-полимераза III).
В составе РНК-полимеразы E.coli выделяют четыре субъединицы: две
α-субъединицы, по одной β- и β’-субъединице. Имеется также дополнительный бел-
ковый σ-фактор Последний необходим только для связывания с промотором и не участвует в удлинении цепи РНК.
Строение РНК-полимераз эукариот имеет много общего со структурой бактери-
ального фермента: они имеют по две больших субъединицы и несколько малых субъединиц.
СТАДИИ ТРАНКРИПЦИИ
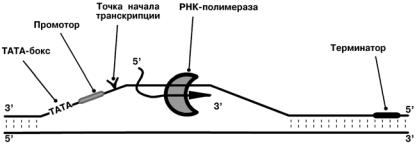
Инициация
Промотор содержит стартовый сигнал транкрипции ТАТА-бокс – определенную
последовательность нуклеотидов ДНК, присоединяющий инициирующий ТАТА-фактор. Этот ТАТА-фактор обеспечивает присоединение РНК-полимеразы к
той нити ДНК, которая будет использоваться в качестве шаблона для транскрипции.
Так как промотор ассиметричен, то он связывает РНК-полимеразу только в одной
ориентации, что определяет направление транскрипции от 5’-конца к 3’-концу
(5’ → 3’).
Другие факторы инициации раскручивают спираль ДНК перед РНК-полимеразой. После синтеза затравочного фрагмента РНК длиной 8-10 рибонуклеотидов
σ-фактор отрывается от фермента.
Элонгация
Белковые факторы элонгации обеспечивают продвижение РНК-полимеразы
вдоль ДНК и расплетание нитей ДНК на протяжении примерно 17 нуклеотидных пар. РНК-полимераза продвигается со скоростью примерно 40-50 нуклеотидов в секунду
в направлении 5’ → 3’. Используя одновременно в качестве субстрата и источника
энергии АТФ, ГТФ, ЦТФ, УТФ.
Терминация
РНК-полимераза остановится, когда достигнет терминирующих кодонов. С по-
мощью белкового фактора терминации, так называемого ρ-фактора (греч. ρ - "ро"),
от матрицы ДНК отделяются фермент и синтезированная молекула РНК, которая
является первичным транскриптом, предшественником мРНК или тРНК или рРНК.

10
ПРОЦЕССИНГ РНК.
Снтезированные молекулы РНК являются и в дальнейшем претерпевают ряд
изменений, которые называются процессингом. У эукариот процессингу подвергаются все виды пре-РНК, у прокариот – только предшественники рРНК и тРНК.
ПРОЦЕССИНГ ПРЕДШЕСТВЕННИКА МРНК
1.Кэпирование (англ. c a p - шапка) – происходит еще во время транскрипции,
состоит в том, что к 5’-трифосфату концевого нуклеотида пре-мРНК присоединяется
5’-углерод N7-метил-гуанозина. «Кэп» необходим для защиты молекулы РНК от
5’-3’-экзонуклеаз.
2. При транскрипции зон ДНК, несущих ин-
формацию о белках,
образуются гетерогенные ядерные РНК, по
размеру намного пре-
восходящие мРНК. Де-
ло в том, что из-за мо-
заичной структуры генов эти гетерогенные
РНК включают в себя
информативные (экзоны) и неинформативные
(интроны) участки. При
особом |
процессе – |
|
сплайсинге |
(англ. |
|
s p l i c e |
– |
склеивать |
встык) происходит удаление интронов и со-
хранение экзонов.