

15.7. Нейронная организация локомоции
Структуры, принимающие участие в организации движений, располагаются во всех отделах мозга. Они взаимосвязаны морфологически или функционально. Наличие связей между центрами организации движений позволяет создавать и реализовывать специальную программу управления движениями. Программа должна быть открытого типа, т.е. позволять изменять, корректировать последующий двигательный акт от результата предыдущего движения. Таким путем создаются условия для саморегуляции двигательного акта.
Регуляция моторных функций по уровням их организации может быть представлена следующим образом.
Спинальный уровень. На этом уровне осуществляется наиболее простая форма автоматического регулирования состояния мышц (эффектора) — рефлекс на растяжение. В основе этого рефлекса лежит обратная связь от мышечных веретен к альфа- мотонейронам спинного мозга. Структурно этот рефлекс состоит из рецептора интра-фузального волокна, афферентного пути через спинальный ганглий, альфа-мотонейрона, экстрафузального волокна. Сила сокращения последнего зависит от силы растяжения интрафузального волокна и от влияния центральных структур на мотонейроны спинного мозга. Мотонейрон может иннервировать от нескольких единиц до 2000 мышечных волокон. Объединение мотонейрона с иннервируемыми им волокнами называется моторной единицей.
Управление работой мышцы осуществляется не отдельным нейроном, а мотонейронным пулом. Мотонейронный пул определяет силу и участие в сокращении всех волокон или их части каждой мышцы. Помимо управления силой сокращения мышцы, мотонейронный пул определяет волокна, которые должны сокращаться в данном движе- нии, и насколько сильно будет это сокращение. Именно этот ме-ханизм позволяет мышце сокращаться не одинаково при разных видах движений. Поскольку в мотонейронном пуле нейроны имеют разные пороги чувствительности к силе раздражения, то при слабом
47
раздражении в сокращении будет принимать участие только часть волокон, при сильном — в сокращении участвуют волокна всей мышцы.
Между мотонейронными пулами мышц сгибателей и разгибателей имеют место реципрокные отношения их активности. Мотонейронный пул является управляющей и управляемой системой. Управление состоянием отдельных нейронов пула осуществляется вышележащими структурами центральной нервной системы.
Таким образом, регуляция локомоций на спинальном уровне начинается с возбуждения рецепторов интрафузальных волокон. От них через задние корешки спинного мозга сигнал о состоянии мышечных волокон поступает -к мотонейронам пула передних рогов спинного мозга. Мотонейроны пула посылают сигнал к мышечным экстрафузальным волокнам своей моторной единицы. Чем сильнее возбуждение рецепторов интрафузальных волокон, тем больше нейронов одного пула возбуждается, и, следовательно, больше экстра-фузальных волокон сокращается, чем достигается сильное сокращение мышцы.
Описанный спинальный уровень организации движений обеспечивает автоматические двигательные реакции. Вышележащие структуры центральной нервной системы управляют деятельностью спинальных центров, причем, эти управляющие влияния могут быть как возбуждающими, так и тормозными.
На рис. 15.7 представлена схема взаимодействия нейронных нервных уровней организации движений. Первый уровень — спинальный, второй — ствола мозга и третий программируемый уровень — высший, корковый. Все три уровня регуляции движений могут ре-ализовывать эти функции как самостоятельно, так и с участием других уровней. Причем, каждый из этих уровней может регулировать работу мышц через мотонейроны спинного мозга как последовательно, так и параллельно. Следовательно, любое сокращение мышцы может быть вызвано спинным мозгом, структурами ствола и коры. Совокупное участие разных уровней позволяет повысить надежность регуляции движения, их точность, локальность, сложность.
Каждый уровень регуляции имеет обратные связи об исполнении движения от мышечной системы, каждый уровень регуляции, посылая команду к мотонейронам спинного мозга, одновременно посылает сигнал о команде к другим выше- или нижележащим центрам. Это позволяет программирующему центру своевременно оценивать команды других уровней и в нужный момент производить коррекцию управления движений.
Стволовые центры регуляции движений представлены нисходящими путями, идущими к спинному мозгу. Ретикулоспинальный путь продолговатого моста, спускаясь к мотонейронам спинного мозга, обеспечивает быстрое перераспределение тонуса мышц при стоянии и ходьбе.
Ретикулярные ядра продолговатого мозга получают информацию от рецепторов органа равновесия и от мозжечка. Нисходящие пути
48

Рис.15.7. Главные отделы головного мозга, участвующие в
прямом управлении движениями по нисходящим путям к мотонейронам.
латерального вестибулярного тракта заканчиваются на мотонейронах, иннервирующих разгибатели конечностей. Раздражение латерального вестибулярного ядра повышает тонус мышц разгибателей, создавая условия поддержания позы.
Медиальное вестибулярное ядро обеспечивает движения глаз, сопряженные с поворотом головы.
Красное ядро среднего мозга при его раздражении приводит к сгибанию конечностей. Причем, мотонейроны мышц сгибателей спинного мозга при раздражении красного ядра возбуждаются, а мотонейроны разгибателей — тормозятся. Следовательно, поскольку возбуждение красного ядра усиливает сгибание конечностей, а раздражение вестибулярного ядра усиливает тонус разгибателей, то эти ядра находятся в реципрокных отношениях. Взаимодействие ретику-лоспинального, вестибулярного и красного ядер имеет определенную последовательность.
Начало движения конечности при шагании сопровождается усилением активности нейронов ретикулоспинального пути, идущего к мотонейронам спинного мозга. Тонус мышц сгибателей повышается, разгибателей — снижается. Затем включаются вестибулярные нейроны, активирующие мотонейроны мышц разгибателей, которые распрямляют конечность и удерживают ее после шага при опоре, про-
49
тиводействуя силе тяжести. В момент отрыва конечности от опоры и начала ее переноса включаются нейроны красного ядра, они координируют работу мотонейронов мышц сгибателей и мотонейронов разгибателей (рис. 15.8).
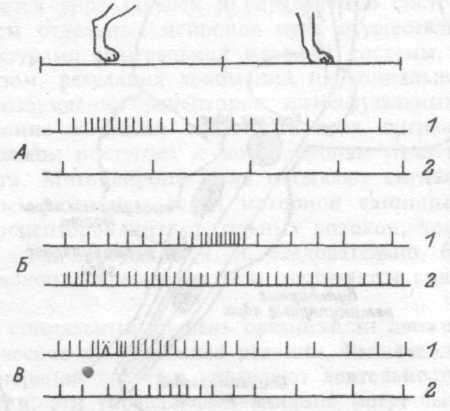
Рис.15.8. Активность одиночных нейронов в двигательных центрах ствола мозга на протяжении цикла перестановки конечности.
1 — до удаления мозжечка, 2 — после удаления; А — ретикупоспинапьный, Б — вестибулярный, В — руброспинальный нейроны.
Регуляция активности указанных трех ядер осуществляется мозжечком и сенсорной обратной связью от проприорецепторов конечностей. Мозжечок создает общую генерализованную активацию ядер, тонизирует их. Сенсорная активация моделирует мозжечковую, приурочивает усиление импульсации нейронов к соответствующему моменту движения.
Произвольные движения человека регулируются корой головного мозга. Составлены подробные карты локализации в коре пунктов организации движений ног, рук, головы, лица, туловища. Показано, что все поля, соответствующие разным частям тела, перекрещиваются, но в них имеются участки с наименьшими порогами, т.е. наиболее чувствительные для вызова определенного вида движения.
Управление мотонейронами спинного мозга при произвольных движениях осуществляется прецентральной бороздой коры (поле 4). Это управление частично реализуется через аксоны клеток Беца, образующих кортикоспинальный путь, оканчивающийся на альфа-мотонейронах спинного мозга. Кортикоспинальный (так называемый, пирамидный) путь только на 3% состоит из аксонов клеток Беиа.
50
Остальная часть этого пути начинается от экстрапирамидной коры поля 6, полей 1, 2, 3. Стимуляция этих полей вызывает движения, но они менее точны, чем при стимуляции поля 4. Параллельно с пирамидными путями к мотонейронам спинного мозга идут экстрапирамидные: ретикуло- , вестибуло- , рубро- , тектоспиналъные пути.
Аксоны пирамидного пути дают коллатерали во всех структурах! ствола мозга, через которые проходят. Следовательно, пирамидный и экстрапирамидный пути образуют взаимные связи.
Реализация произвольных движений с участием перечисленных структур пирамидной и экстрапирамидной систем осуществляется следующим образом. Для начала любого произвольного движения необходимо провести выбор мышц, с помощью которых это движение может быть осуществлено. Этот выбор обусловлен исходной позой, положением конечностей, о чем информируется экстрапирамидная и проприоцептивная системы, производящие оценку тонуса мускулатуры.
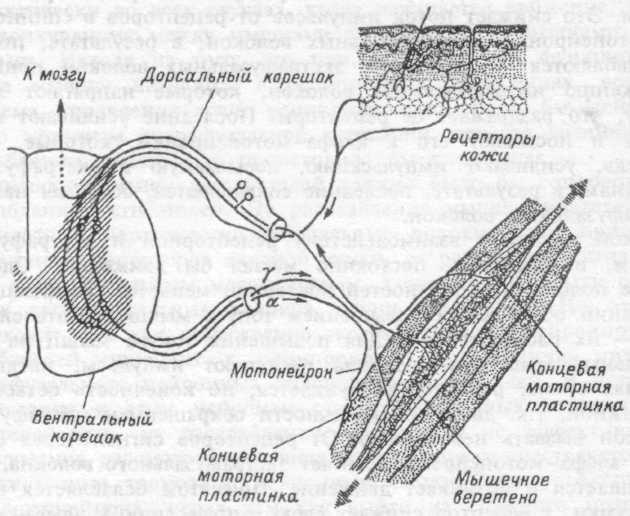
Рис.15.9. Обшая схема системы гамма-мотонейрона.
Мышца содержит несколько сотен двигательных единиц. Мотонейроны, иннервируюшие одну мышцу (мотонейронный пул), располагаются в нескольких сегментах спинного мозга. Аксоны мотонейронов и сами мотонейроны не однотипны. Около 30% аксонов являются тонкими волокнами, они по классификации относятся к группе гамма волокон, соответственно, нейроны, которым они принадлежат, называют гамма-мотонейронами (рис. 15.9).
57
Мышечные веретена имеются во всей скелетной мышце. В каждом мышечном веретене содержится по 2-10 мышечных волокон. В центральной части веретена находится эластичная ядерная сумка, в ней расположены рецепторы, в том числе так называемые аннулос-пинальные рецепторы. Интрафузальные мышечные волокна одним концом крепятся к ядерной сумке, другим — к сухожилию или оболочке экстрафузального волокна.
Рецепторы ядерной сумки раздражаются тремя способами. При сокращении экстрафузальных волокон они уменьшаются, при этом натяжение мышечного вертена также уменьшается. Когда мышца расслабляется, веретено натягивается, удлиняются интрафузальные волокна, они растягивают сумку. Это раздражает расположенные в ней рецепторы. Второй механизм раздражения сумки — сокращение интрафузальных волокон. Третий механизм — внешняя механическая нагрузка на мышцу, удлиняющая ее, одновременно растягивающая ядерную сумку и раздражающая ее рецептор.
От рецепторов ядерной сумки афферентный нерв идет в тот сегмент, где находится мотонейронный пул длинной мышцы и посылает аксоны к экстрафузальным волокнам. Таким образом, экстра-фузальное волокно механически связано с веретеном, а от веретена к экстрафузальному волокну идет нервная связь: рецептор сумки -> афферентный нерв -> сегмент спинного мозга -> альфа-мотонейрон -> экстрафузальное волокно.
Сокращение экстрафузального волокна ослабляет натяжение интрафузальных, а затем уменьшает раздражение рецепторов ядерной сумки. Это снижает поток импульсов от рецепторов в спинной мозг, к мотонейронам экстрафузальных волокон, в результате, последние расслабляются. Расслабление экстрафузальных волокон приводит к натяжению интрафузальных волокон, которые напрягают ядерную сумку, что раздражает ее рецепторы. Последние усиливают возбуждение и посылают его к альфа-мотонейронам, которые, в свою очередь, усиливают импульсацию, посылаемую к экстрафузальным волокнам, в результате, последние сокращаются, ослабляя натяжение интрафузальных волокон.
Такой механизм взаимодействия рецепторной и экстрафузальной систем, в принципе, постоянно мешал бы движению. Однако, в норме положение конечностей постоянно меняется. Реализация этих движений обусловлена повышением тонуса мышц сгибателей, а затем — их расслаблением. Для повышения тонуса мышц на интрафузальные мышечные волокна поступают импульсы, натягивается ядерная сумка, рецептор возбуждается, но конечность остается неподвижной, т.к. движение конечности сокращением интрафузальных волокон вызвать невозможно. От рецепторов сигнал через спинной мозг, альфа-мотонейрон, достигает экстрафузального волокна, мышца сокращается и возникает движение. При этом ослабляется натяжение сумки, а рецептор снижает свою импульсацию к альфа-клеткам. В итоге, конечность принимает новое положение, которое сохраняется, пока не изменится активность на входе интрафузальной системы.
52
Обратную связь от интрафузальных волокон к экстрафузальным мотонейронам иллюстрирует сухожильный коленный рефлекс. При нанесении удара по сухожилию подколенной чашечки мышца натягивается, рецепторы ее веретен раздражаются и посылают возбуждение к мотонейронному пулу экстензоров, что и вызывает сокращение мышцы.
Интрафузальные нейроны, интрафузальные волокна и обратная связь от последних являются решающими в организации позных рефлексов типа Магнуса-Клейна. При этих рефлексах, когда голова животного отводится назад — передние конечности разгибаются, задние —сгибаются. При наклоне головы вперед — передние конечности сгибаются, задние — разгибаются. При повороте головы происходит разгибание конечностей на стороне, куда повернута голова и сгибание конечностей на противоположной. Эти рефлексы сохраняются даже у децеребрированного животного, что свидетельствует о их реализации на уровне спинного мозга.
Точно также, за счет воздействия на интрафузальные волокна запускаются рефлексы отдергивания при болевых раздражениях, че-сательный рефлекс, произвольные двигательные реакции.
Параллельно с механизмом регуляции движений через интрафу-зальную систему сушествует система регуляции, которая запускается аппаратами Гольджи — рецепторов сухожилий. Сигнал от этих рецепторов тормозит активность мотонейронного пула, когда натяжение сухожилий становится чрезвычайно сильным. Эта рецепторная система защищает мышцу от перенапряжений.
Практически во всех случаях, когда начинается движение, возникает конкуренция между мышцами сгибателями и мышцами разгибателями. Исходя из того, что при движении эти функционально разные мышцы находятся в реципрокных отношениях, возникает проблема управления этой реципрокностью. Если бы действовал только механизм интрафузальной активации, которая возникает при расслаблении мышцы, то происходило бы следующее.
Сокращение мышц разгибателей должно было бы сопровождаться расслаблением сгибателей. Но расслабление мышц сгибателей будет натягивать в веретене интрафузальные волокна, это приведет к раздражению рецепторов ядерной сумки, от рецепторов сигнал пойдет к экстрафузальным мотонейронам, а они должны были бы возбудить экстрафузальные волокна мышц сгибателей. Но этого не происходит, так как возбуждение экстрафузального нейронного пула разгибателей приводит к реципрокному торможению интра- и экстрафузального нейронных пулов мышц сгибателей. В результате, импульсация к этим нейронам от интрафузальных волокон мышц сгибателей при их расслаблении оказывается не эффективной.
Реализация рассмотренного вида торможения осуществляется через промежуточные тормозные нейронные клетки Реншоу. Аксон каждого мотонейрона, перед тем как выйти из спинного мозга, дает коллатерали, заканчивающиеся на тормозных клетках Реншоу. Последние посылают аксон на клетки тех мотонейронов, которые прислали к нему свои коллатерали и на клетки, находящиеся с ними
53
в реципрокных отношениях. Так формируется отрицательная обратная тормозная связь или возвратное торможение. Спинальный уровень регуляции моторики находится в зависимости от вышележащих структур центральной нервной системы. Эта зависимость определяется пирамидной и экстрапирамидной системами.
Аксоны нейронов моторной коры идут в составе внутренней капсулы, затем в мост. 80% этих аксонов перекрещиваются на уровне продолговатого мозга и идут к вставочным нейронам задних рогов. 20% аксонов пирамидного тракта, неперекрещиваясь, спускаются к мотонейронным пулам, преимущественно заканчиваясь на промежуточных нейронах этих пулов.
Стимуляция моторной коры, и особенно вторичной моторной зоны, вызывает целостное движение, а не подергивание отдельных мышц. Повреждения моторной коры пирамидного тракта у человека, возникающие в результате травм, кровоизлияний, приводят к потере мышечного тонуса (вялый паралич), утрачивается способность выполнять некоторые виды движений, хотя мышцы могут участвовать в других движениях. Например, при кровоизлиянии в определенную зону коры больной не может согнуть отдельно палец, но может сжимать и разжимать кулак.
Повреждение двигательной коры за счет утраты тормозного влияния на экстрапирамидную систему, на спинальные рефлексы вызывает при восстановлении функций нижележащих структур нарушения в виде гиперрефлексии, гипертонуса мышц, т.е. после вялого паралича развивается спастический.