

Глава 17. Функции головного мозга
Головной мозг — передний отдел центральной нервной системы, обеспечивающий регуляцию жизненных функций организма, материальный субстрат высшей нервной деятельности (поведения). Он регулирует дыхание, кровяное давление и температуру, положение тела, движения, рефлексы, еду и питье, гормональный статус — почти все вегетативные функции организма. С помощью сенсорных систем головной мозг обеспечивает взаимодействие организма с внешним миром и осуществляет контроль внутренней среды. Высшие функции мозга, которые рассматриваются в данной главе, связаны с умственной деятельностью человека — это сенсорное восприятие и целенаправленное движение, обучение и память, эмоции, речь и мышление, бодрствование и сон, и, наконец, сознание, как осознание умственной и (или) физической деятельности.
Конечным результатом восприятия раздражителя является организация эффекторного управляющего сигнала, реализующегося, как правило, в движении. Специфические сенсорные системы перерабатывают информацию о раздражителях, после чего она поступает к различным исполнительным структурам мозга. Двигательная система, также как и сенсорные системы, обладает многоуровневой организацией и обеспечивает последовательную переработку управляющего движением сигнала: от моторной коры до мышечного сокращения по команде спинальных мотонейронов. Хотя сенсорные и двигательная системы, в принципе, независимы друг от друга и могут работать изолированно, очень редко ощущение не сопровождается двигательной реакцией. Неразрывная связь ощущения и движения четко определена И.М.Сеченовым, который в "Рефлексах головного мозга" (1863) показал, что без внешнего чувственного раздражения невозможна психическая деятельность и ее объективное выражение — мышечное движение.
Движения бывают непроизвольными — рефлекторными и произвольными — целенаправленными. Непроизвольные движения входят в систему обеспечения безусловных рефлексов, непроизвольные и произвольные движения совместно обеспечивают условные рефлексы, вырабатываемые на различные раздражители.
17.1. Безусловные и условные рефлексы
Безусловные рефлексы — индивидуальные и видовые, генетически закрепленные, осуществляемые с участием центральной нервной системы стереотипные реакции организма на внешние и внутренние
114
раздражители (стимулы). К безусловным рефлексам относятся, например, выделение желудочного сока при попадании пищи в рот, отдергивание руки при болевом раздражении, питье жидкости при изменении кислотно-щелочного равновесия в организме, мигание при попадании воздуха в глаз и т.д. В таблице 17.1 представлены безусловные рефлексы различной степени сложности.
Таблица 17.1 Безусловные рефлексы
|
Тип рефлекса |
Название по функции |
Назначение |
Структуры мозга, раздражение или разрушение которых заметно влияет на рефлекс |
|
Индивидуальные рефлексы |
Питьевой Пищевой Агрессивный Оборонительный |
Рефлексы самосохранения |
Латеральный и медиальный гипоталамус, гипофиз, центральное серое вещество вокруг сильвиева водопровода, миндалина |
|
|
Исследовательский Игровой Имитационный Свободы (преодоления) |
Рефлексы саморазвития |
Ядра миндалевидного комплекса, гиппокамп, лобная кора |
|
Видовые рефлексы |
Половой Родительский Территориальный Иерархический |
Рефлексы сохранения вида — ролевые, социальные |
Обонятельная кора, лобная кора, перегородка, гиппокамп |
И.П.Павлов — основоположник объективного изучения функций головного мозга с помощью разработанного им метода условных рефлексов — физиологии высшей нервной деятельности (поведения). Условные рефлексы (временный рефлекс, временная связь, условная связь, классическое обусловливание) — индивидуально приобретенные в процессах жизни или специального обучения системные приспособительные реакции, возникающие на основе образования временной связи между условным (сигнальным) раздражителем и без-условнорефлекторным актом. Термины "условный рефлекс" и "безусловный рефлекс" были предложены И.П.Павловым (1903).
Условный рефлекс образуется благодаря многократному сочетанию индифферентного (относительно вырабатываемой реакции) раздражителя со стимулом, вызывающим безусловный рефлекс. Например, многократное включение звонка, предшествующего пище, приводит к выделению слюны и желудочного сока у животного только на звонок. При этом звонок становится условным раздражителем или условным стимулом (сигналом), подготавливающим организм к пищевой реакции. Между стимулом и реакцией в процессе образования условного рефлекса формируется функциональная связь. Условный рефлекс приобретается в процессе обучения, причем, функциональная связь между рецепторами, на которые действует условный
115
раздражитель, и эффекторами, обеспечивающими условнорефлектор-ный ответ, формируется на основе совпадения условного раздражителя с безусловным рефлексом.
Безусловные раздражители, вызывающие безусловные рефлексы, базируются на врожденных фиксированных связях между рецепторами и эффекторами. Структурной предпосылкой формирования условнореф-лекторных реакций является конвергенция условного и безусловного раздражителя на одних и тех же нейронах головного мозга.
Связь между стимулом и реакцией: классический условный рефлекс. Выработка классического условного секреторного рефлекса начинается с того, что сначала "запускается" безусловный рефлекс, например, на получение пищи у животного выделяется слюна. Затем стимул, вызывающий безусловный рефлекс (пища), многократно сочетают с произвольно выбранным любым другим индифферентным стимулом — звуком, светом, прикосновением, электрическим раздражением. Через некоторое число сочетаний слюна начинает выделяться уже не на пищу, а на изначально индифферентный стимул, который в процессе обучения становится условным сигналом. Наступает стадия, когда условный рефлекс сформировался. Это значит, что уже возникла и упрочилась ассоциация между условным стимулом и безусловным стимулом, адекватным для запуска безусловного рефлекса. Некоторые условные рефлексы перечислены в таблице 17.2.
Для выработки условного рефлекса необходимо соблюдение трех правил. Во-первых, условный (первоначально индифферентный) раз-
Таблица 17.2 Условные рефлексы
(классические и инструментальные)
|
№ |
Наименование |
Способ выработки |
Примечания |
|
п/п |
рефлекса |
|
|
|
1 |
Первого порядка |
На сочетание индифферентного раздра- |
Простые формы реф- |
|
|
|
жителя с безусловным. |
лексов — пищевые, |
|
2 |
Натуральный |
На раздражитель, обладающий есте- |
оборонительные и т.д. |
|
|
|
ственными свойствами безусловного |
|
|
|
|
раздражителя (например, запах или вид |
|
|
|
|
пищи). |
|
|
3 |
На время |
На определенный промежуток времени |
Разновидности услов- |
|
|
|
между условным и безусловным раз- |
ного рефлекса, при |
|
|
|
дражителем. |
котором подкрепление |
|
4 |
Следовой |
На любой условный стимул, через не- |
применяется после |
|
|
(отставленный ) |
которое время после прекращения ко- |
завершения действия |
|
5 6 |
Наличный (отставленный ) Запаздывающий |
торого, применяется подкрепление. На время между условным и безусловным раздражителем, включающимся на фоне условного через 5-30 с. На время при длительном периоде изолированного действия условного раздражителя, к которому присоединяется условный через 30-180 с. |
условного стимула. Разновидности условного рефлекса, при котором подкрепление применяется во время действия условного стимула. |
116

Таблица
17.2 (продолжение)
|
№ Наименование п/п рефлекса |
Способ выработки |
Примечания |
|
7 Коротко-отставленный (совпадающий) 8 На комплексные раздражители 9 На цепь раздражителей 10 На отношение раздражителей 11 Высшего порядка |
На условный сигнал через 5 с, после которого дают подкрепление. На одновременный комплекс раздражителей или на последовательность раздражителей. На последовательные, разобщенные некоторой паузой, раздражители. На относительные, а не на абсолютные, свойства раздражителей. Образованные на базе другого условного рефлекса. |
Сложные формы раздражителей. Выработка рефлексов на такие раздражители требует участия корковых зон: проекционных и ассоциативных. Лежат в основе организации поведения. Требуют участия проекционных, ассоциативных и лобных (лимбических) зон. |
дражитель
должен предварять действие безусловного
стимула. Например,
при выработке пищевого условного
рефлекса на звук необходимо,
чтобы звук включался раньше появления
пищи. Через несколько
сочетаний действия звука и следующего
за ним пищевого подкрепления
слюна и желудочный сок будет выделяться
при действии
звука, независимо от последующего
получения пищи. Во-вторых,
биологическая
значимость условного раздражителя.
должна быть
меньше, чем безусловного. Например, для
кормящей матери крик
ребенка будет более сильным раздражителем,
чем пищевое подкрепление.
Поэтому выработка пищевого условного
рефлекса у матери
на крик ребенка — звук столь высокой
биологической зна-чимости-не
будет успешной. В-третьих,
сила
как условного, так и безусловного
раздражителей должна быть определенной
величины (закон
силы): слишком слабые и слишком сильные
раздражители не позволяют
выработать стабильный условный рефлекс.
После стабилизации и закрепления многие условные рефлексы становятся автоматическими действиями, например, нажатие водителем автомобиля на тормоз при появлении препятствия. Если условный раздражитель не подкрепляется, условный рефлекс угасает.
Классический условный рефлекс, выработанный на сочетание условного сигнала и безусловного подкрепления, называется условным рефлексом первого порядка. Условный рефлекс, образованный на базе другого условного рефлекса, называется условным рефлексом второго порядка. На его основе можно выработать рефлекс третьего порядка и т.д.
Механизмы условных рефлексов изучались на животных морфо-функциональными, фармакологическими и электрофизиологическими методами, а также с помощью экстирпаций различных участков мозга
117
 или
перерезок проводящих путей. И.П.Павлов
считал, что формирование
временных связей (условный сигнал —
безусловное подкрепление)
является функцией коры больших полушарий
мозга, поскольку
животные, лишенные проекционных зон
коры вырабатывают
только очень элементарные условные
рефлексы. В дальнейшем, установили,
что наряду с корковыми отделами мозга
в условнореф-лекторных
реакциях принимают участие ретикулярная
формация и лимбическая
система.
или
перерезок проводящих путей. И.П.Павлов
считал, что формирование
временных связей (условный сигнал —
безусловное подкрепление)
является функцией коры больших полушарий
мозга, поскольку
животные, лишенные проекционных зон
коры вырабатывают
только очень элементарные условные
рефлексы. В дальнейшем, установили,
что наряду с корковыми отделами мозга
в условнореф-лекторных
реакциях принимают участие ретикулярная
формация и лимбическая
система.
Генерализованные, широко распространенные неспецифические формы активности нейронов при выработке условных рефлексов выявлены в таламусе, стриопаллидарной системе, мозжечке и гипоталамусе. Это значит, что в процессе условнорефлекторного обучения принимают участие многие структуры головного мозга. По мере упрочения условного рефлекса генерализация активности различных структур мозга уменьшается. На конечной стадии формирования стабильного условного рефлекса в этот процесс вовлекаются нейроны только тех областей мозга, которые имеют непосредственное отношение к анализу условного и безусловного раздражителя и к осуществлению условнорефлекторной реакции. В моторной коре, управляющей движением, а также в корковых представительствах безусловных рефлексов возникают специфические реакции нейронов, предваряющие условнорефлекторный ответ.
При выработке условного рефлекса усиливается активация мозга, источником которой являются десинхронизирующие влияния, исходящие из ретикулярной формации. В период выработки пищевых и оборонительных рефлексов у нейронов головного мозга, вовлеченных в реакцию, частота разрядов возрастает от сочетания к сочетанию условного и безусловного раздражителя. Так происходит до достижения 60% уровня выполнения условного рефлекса. После достижения этого уровня активность нейронов то увеличивается, то уменьшается, т.е. носит волнообразный характер. На этой стадии условный раздражитель вызывает стабильный для многих нейронов кратковременный ответ, предваряющий условнорефлекторную реакцию.
Для характеристики механизма условного рефлекса надо знать нейрофизиологические, биофизические и биохимические изменения, происходящие при ассоциативном (условнорефлекторном) обучении. Нейрофизиологические изменения выявлены при изучении мозга животных с помощью микроэлектродов. Это изменения возбудимости определенных нейронов в процессе обучения. Биофизические изменения при выработке условных рефлексов изучались только на бес-позвоночных животных. Оказалось, что ионы кальция аккумулируются в некоторой части клеток (в основном, клеток тормозного типа), благодаря снижению количества калиевых каналов, что увеличивает возбудимость клетки. Биохимические изменения связаны с фосфорилированием, процессом, который меняет свойства белка ионных каналов в клеточной мембране и, соответственно, ведет к изменению функциональной активности ионных каналов. Действие фосфорилирующих ферментов сохраняется длительное время после завершения обучения. Данные, полученные на низкоорганизованных
118
 животных,
еще не достаточны для ответа на вопрос,
происходят ли сходные
процессы у высших животных и человека.
Возможно, что клеточные
механизмы обучения сохранились в
эволюции, но дополнились
в сложных системах механизмами длительного
хранения следов
раздражителей.
животных,
еще не достаточны для ответа на вопрос,
происходят ли сходные
процессы у высших животных и человека.
Возможно, что клеточные
механизмы обучения сохранились в
эволюции, но дополнились
в сложных системах механизмами длительного
хранения следов
раздражителей.
Большое значение в организации поведения играет ориентировочный рефлекс, описанный И.П.Павловым под названием рефлекс "что такое?". Он возникает в ответ на неожиданные и новые раздражители, но участвует и при их повторении, постепенно угасая. Величина и длительность сохранения ориентировочного рефлекса зависит от физических и биологических свойств раздражителя и "нервной модели" сигналов того же класса, сформированной ранее и сохраняемой в памяти субъекта. Ориентировочный рефлекс является двухфазным процессом: первая фаза — фаза неспецифической тревоги, вторая — исследовательского поведения. При реализации этого рефлекса включается движение глаз в направлении стимула, кожно-гальванический рефлекс, сосудистые реакции, наблюдается депрессия альфа-ритма. Вегетативные компоненты рефлекса исчезают первыми (обычно после 10-12 предъявлений стимула), позже исчезают сосудистые реакции и депрессия альфа-ритма. Восстановление ориентировочного рефлекса возможно только при изменении физических параметров и биологической значимости раздражителя. Считают, что афферентный стимул активирует ретикулярную формацию ствола мозга и кору мозга по двум параллельным путям — специфическому (лемнисковому) и неспецифическому (экстралемнисковому). Такая активация обуславливает первую фазу ориентировочной реакции. Вовлечение после первичной активации коры также лимбических структур, тормозящих нейроны ретикулярной формации, создает условия для реализации второй фазы ориентировочной реакции — исследовательского поведения.
Значение ориентировочной реакции в организации поведения связано также с образованием ассоциативной связи по типу условного рефлекса между двумя индифферентными раздражителями. Так, постоянное сочетание звука и света после упрочения связи между этими раздражителями обуславливает на звук ту же реакцию, что и на свет. Например, сильное расширение зрачка, вызванное светом, при многократном сочетании со звуком может возникать при действии только звука.
Изучение условных рефлексов необходимо для получения данных о том, как приобретаются сведения о связях между раздражителями и реакциями организма. Однако, условнорефлекторное обучение и поведение не исчерпываются лишь классическими условными рефлексами. Связь между реакцией и подкреплением — инструментальный условный рефлекс.
Инструментальный условный рефлекс — это условный рефлекс, при котором реакция на условный раздражитель (обычно двигательная) является обязательным условием получения подкрепления. Например, если нажатие на педаль, сопровождаемое получением жи-
119


вотным
пищи, предваряется звуковым или световым
раздражителем, то
после ряда сочетаний нажатие на педаль
становится инструментальной
реакцией, а
внешний стимул — сигналом для совершения
такой
реакции. Это и есть инструментальный
условный рефлекс. Такой
вид обучения, основанный на активной
деятельности, играет важную
роль в организации поведения человека
в раннем постна-тальном
онтогенезе и остается доминирующим на
протяжении дальнейшей
жизни. Примером инструментальной реакции
ребенка является
его плач, в результате которого ребенок
получает пищу. В этом случае
плач играет для ребенка роль инструмента,
изменяющего условия
среды. Ребенок овладевает реакцией,
которая определяет получение
поощрения.
Инструментальный условный рефлекс успешно вырабатывается не только на поощрение, но и на наказание. Так, если собаку поместить в помещение, разделенное барьером, с целью научить ее перепрыгивать через него, для эффективного обучения возможно использование не поощрения (пища), а наказания (ток). Металлический пол подвергается воздействию электрического тока и животное, испытывая неприятное ощущение или боль, быстро обучается преодолевать препятствие и оказываться за барьером, где пол не находится под напряжением. При этом, одновременно с током включается световой или звуковой сигнал. В дальнейшем, если за 10 секунд до включения тока загорается световой или слышен звуковой сигнал, то собака избегает "наказания" — она перепрыгивает через барьер в течение того времени, которое отделяет световой или звуковой сигнал от включения тока.
Инструментальные условные рефлексы отличаются от классических условных рефлексов тем, что на их основе возникает бесконечное разнообразие новых двигательных реакций. Таким образом, поведение формируется на основе результата действия. Закрепляются в памяти субъекта в этом случае не безусловно-рефлекторные реакции, а произвольные активные действия.
Подкрепление (например, пища, электрический ток) является средством, помогающим человеку или животному обучиться той реакции, за которой это подкрепление следует. В то же время, подкрепление в инструментальном обучении служит сигналом, который указывает на то, какие действия желательны, целесообразны, а какие — нет. Максимальную эффективность подкрепление достигает при условии его воздействия непосредственно после совершения действия (порядка 5 секунд). Подкрепление может исходить из внешнего источника — например, получение пищи за нажатие на определенный рычаг, или из внутреннего источника, например, возникновение положительной эмоции. Известны эксперименты с вживлением в мозг крысы электродов, по которым животное, путем нажатия на рычаг, замыкающий электрическую цепь, могло посылать к различным структурам импульсы электрического тока. Подобное раздражение ряда лимбических структур мозга, так называемых, "центров удовольствия", осуществлялось крысой непрерывно, до полного изнеможения. Это — пример инструментального обучения с внутренним подкреплением — получением положительной эмоции.
120


 В
механизмах реализации инструментального
условного рефлекса наиболее
выраженное участие принимают нейроны
двигательной коры
и центрального серого вещества,
окружающего сильвиев водопровод.
Реакции нейронов этих отделов мозга
возникают на 500-800 мс
раньше начала движения. Условный
раздражитель (например, звуковой
щелчок) вызывает в тех же нейронах
длительный тонический
разряд, постепенно угасающий при
отсутствии подкрепления. В выборе
правильной инструментальной реакции,
т.е. движения с последующим
положительным подкреплением, принимают
участие нейроны
лобной коры, в которых импульсация
значительно учащается.
При ошибочном выборе активность нейронов
лобной коры не усиливается
по сравнению с фоном. Нейроны двигательных
ядер таламуса
и пирамидной системы непосредственно
связаны с организацией
движения при инструментальном условном
рефлексе.
В
механизмах реализации инструментального
условного рефлекса наиболее
выраженное участие принимают нейроны
двигательной коры
и центрального серого вещества,
окружающего сильвиев водопровод.
Реакции нейронов этих отделов мозга
возникают на 500-800 мс
раньше начала движения. Условный
раздражитель (например, звуковой
щелчок) вызывает в тех же нейронах
длительный тонический
разряд, постепенно угасающий при
отсутствии подкрепления. В выборе
правильной инструментальной реакции,
т.е. движения с последующим
положительным подкреплением, принимают
участие нейроны
лобной коры, в которых импульсация
значительно учащается.
При ошибочном выборе активность нейронов
лобной коры не усиливается
по сравнению с фоном. Нейроны двигательных
ядер таламуса
и пирамидной системы непосредственно
связаны с организацией
движения при инструментальном условном
рефлексе.
В условнорефлекторном обучении и основанном на нем поведении существенную роль играют следующие факторы: эффективность раздражителя, доминирующая мотивация, избирательность реагирования. Наиболее эффективными являются биологически значимые сигналы, а также первоначально индифферентные раздражители, физические параметры которых соответствуют максимальной чувствительности органов чувств. Наибольший успех в обучении достигается при согласовании мотива и результата, как, в частности, в случае успеха пищевого подкрепления в условиях голода. Избирательность реагирования — конечного звена рефлекторной деятельности, проявляется как во врожденных формах поведения (типа таксисов у животных), так и в приобретенных: например, рефлекс на сгибание вырабатывается легче, чем рефлекс на разгибание конечностей.
Выработка условного рефлекса проходит две стадии. Первая стадия — генерализации — характеризуется тем, что все близкие по физическим параметрам условные раздражители вызывают одинаковую условную реакцию. Вторая стадия — специализации — характеризуется тем, что постепенно в процессе выработки условного рефлекса различные раздражители, кроме условного, становятся неэффективными и не вызывают условного рефлекса.
Торможение — механизм формирования условных рефлексов. В
основе стадии специализации условного рефлекса лежит торможение. Торможение делят на два типа: 1) врожденное, безусловное, внешнее торможение; 2) приобретенное индивидуально, условное, внутреннее торможение. Внешнее торможение вызывается посторонним для данной условной реакции внешним стимулом и всегда начинается с ориентировочной реакции. Нейроны двигательной коры реагируют на внешние отвлекающие стимулы так же, как и на условные раздражения. Внешнее торможение вызывает безусловную ориентировочную реакцию и затормаживает условно-рефлекторную реакцию. Это осуществляется в результате включения в безусловную ориентировочную реакцию тех же нейронов коры, которые реагируют и на условный раздражитель.
121
В
зависимости от способа выработки
условного рефлекса описывают
разные виды внутреннего
торможения: угасательное,
диффе-ренцировочное,
запаздывательное и условный тормоз.
Угасательное
торможение
возникает после отмены подкрепления:
условный рефлекс
без подкрепления постепенно угасает.
Дифференцировочное
торможение
вырабатывается на основе положительного
условного рефлекса,
когда в эксперимент вводится раздражение,
близкое по параметрам
к условному, но не сопровождаемое
подкреплением. Запаздывательное
торможение
возникает при постепенном отставлении
во времени
безусловного подкрепления, скрытый
период появления условного
рефлекса увеличивается — рефлекс
запаздывает. Условный
тормоз
вырабатывается
при попеременном предъявлении
подкрепляемого
и неподкрепляемого условного раздражения,
причем последнему
предшествует добавочное раздражение.
Именно добавочное раздражение
приобретает способность тормозить
следующий за ним условный
раздражитель и становится условным
тормозом.
Все эти виды торможения выявляются с помощью экспериментальной процедуры, но основываются на реальных нейрофизиологических процессах. Например, исследование угасательного торможения на уровне одиночных нейронов мозга крыс при пищевом условном рефлексе показало, что самое быстрое угасание реакций при действии неподкрепляемых раздражителей наблюдается у нейронов ретикулярной формации. Реакции нейронов гиппокампа в тех же условиях угасают лишь частично, так же как и реакции нейронов гипоталамуса и преоптической области. Реакции корковых нейронов угасают медленно. Дифференцировочное торможение на уровне одиночных нейронов моторной коры развивается, в основном, в течение первых 5-7 проб, включающих дифференцировочный сигнал. Сначала нейроны реагируют одинаково как на условный, так и на дифференцировочный раздражители. При повторном тестировании появляются и нарастают различия в реакциях на эти раздражители и на конечном этапе выработки дифференцировки реакция на условное положительное раздражение превышает реакцию на дифференцировочное.
Все характерные для угашения и дифференцировки изменения в импульсной активности происходят в тех же структурах мозга, которые принимают участие в выработке условнорефлекторных реакций. Направление перестройки импульсной активности при внутреннем торможении противоположно тому, что наблюдается при выработке условного рефлекса. Внутреннее торможение с нейрофизиологической точки зрения — средство, блокирующее или изменяющее течение приобретенных индивидуальных поведенческих реакций.
Важным средством формирования условнорефлекторной деятельности является доминанта (А.А.Ухтомский), которая представляет собой временно господствующую рефлекторную систему, которая направляет работу нервных центров в данный момент. Если тот или иной центр мозга подготовлен к реакции, благодаря предварительным слабым влияниям внешних или внутренних раздражителей, т.е.
122
возбудимость
его повышена, то возбуждение нарастает
в нем под влиянием
импульсов, поступающих в центральную
нервную систему, независимо
от места и модальности раздражителя.
Многократно усиленное
в доминантном центре возбуждение
тормозит все прочие рефлексы.
Структурно-функционально доминанта
представляет собой
объединение нервных центров, состоящее
из корковых, подкорковых,
вегетативных и гуморальных систем.
Доминанта — не топографически
единый пункт возбуждения в центральной
нервной системе, а синхронно работающее
объединение центров с оптимальным для
данной реакции стационарным возбуждением
в разных этажах головного
и спинного мозга и вегетативной нервной
системе. Взаимодействие
доминанты и условного рефлекса объясняет
схема на рис.17.1.
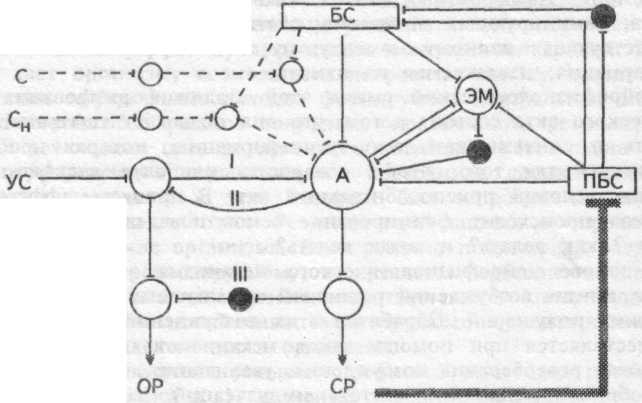
Рис.17.1. Схема взаимодействия доминанты (прерывистые линии) и условного рефлекса (сплошные линии). I — прямая условная связь, II — обратная связь — активирующая, III — тормозящая. Черные кружки — тормозные элементы. Тонкая пиния — влияние подкрепляющего стимула, заштрихованная — инструментальный условный рефлекс. С и Си — индифферентные стимулы; УС — условный стимул; БС — безусловный стимул, запускающий реакцию; ПБС — подкрепляющий безусловный стимул; А — афферентные дуги рефлексов; Э — эфферентные элементы рефлексов; ОР — ориентировочная реакция; СР — специфическая рефлекторная реакция, ЭМ — эмоция.
123
Функциональная
архитектура условного рефлекса. Детальное
изучение
условнорефлекторной деятельности с
использованием электрофизиологических
данных в сопоставлении с вегетативными
и двигательными
компонентами условных реакций, привели
к формулировке
теории
функциональной системы (П.К.Анохин)
как основы понимания
функциональной структуры условного
рефлекса.
Поведение с позиции теории функциональных систем рассматривается как приспособительный акт любой степени сложности, в основе которого лежат следующие процессы: 1) афферентный синтез, 2) стадия принятия решения, 3) формирование акцептора результата действия, 4) формирование интеграла эфферентных возбуждений (эфферентный синтез), 5) получение полезного результата системы, 6) обратная афферентация о параметрах полученного реального результата, сопоставление их с ранее сформировавшимся акцептором результата действия.
Афферентный синтез — это первая стадия поведенческого акта, на которой решается вопрос: "Какой результат должен быть получен системой?". Афферентный синтез включает четыре основные компонента: доминирующая мотивация; обстановочная афферентация, соответствующая данному моменту; пусковая афферентация, а также информация, извлекаемая из памяти.
Нейрофизиологический смысл этой стадии формирования поведенческого акта состоит в том, что она позволяет тщательно обработать и синтезировать всю ту информацию, которая необходима организму для того, чтобы совершить наиболее адекватный для данных условий приспособительный акт. В процессе афферентного синтеза происходит формирование основ поведенческого акта: что делать? как делать? и когда делать?
В основе нейрофизиологического механизма этой стадии лежит конвергенция возбуждений различной модальности на нейронах коры больших полушарий. Обработка этих возбуждений нейронами коры осуществляется при помощи таких механизмов, как корково-подкорковая реверберация возбуждения, увеличение дискриминационной способности нейрона к частоте импульсаций, а также увеличение конвергентной емкости отдельного нейрона с обязательным участием ориентировочно- исследовательской реакции.
Стадия принятия решения является, по существу, стадией, на которой осуществляется формирование конкретной конечной цели, к которой стремится организм. Принятие решения является именно тем критическим пунктом, который переводит один системный процесс — афферентный синтез в другой системный процесс — в программу действий, и является тем переходным пунктом, после которого все комбинации возбуждений приобретают исполнительный характер. Проблема принятия решения на нейрональном уровне состоит из двух частей: 1) задача отдельного нейрона, 2) интеграция нейронов в единую систему.
Одним из наиболее интересных и сложных элементов функциональной системы является акцептор результата действия. Он обеспечивает прогнозирование признаков будущего результата и
124

сравнение
их при помощи обратной афферентации с
параметрами реального
результата системы.
Представление об акцепторе основано на опытах, проводившихся по секреторно-двигательному методу условных реакций активного выбора со стороны подкрепления, на станке с двумя кормушками при подмене подкрепления. Этот прием "сюрпризной ситуации" с подменой подкрепления вызывал у животного бурную ориентировочную реакцию. Значит, в центральной нервной системе еще до получения животным реального результата, к которому оно стремится, формируется "образ" этого результата, а, следовательно, должен существовать и аппарат "подгонки" полученного результата и его прообраза.
Функционирование акцептора результата действия изменяет устоявшиеся представления о работе центральной нервной системы по рефлекторному принципу. Формируясь на основе многостороннего механизма афферентного синтеза, акцептор результата действия отражает непоследовательное развитие явлений поведенческого акта. Он предвосхищает свойства того результата, который должен быть получен в соответствии с принятым решением, т.е. опережает ход событий в отношении между организмом и внутренней средой. Этот аппарат дает организму возможность исправить ошибку поведения.
На стадии эфферентного синтеза формируются центральные механизмы, которые обеспечивают получение определенного результата.
В ходе реализации целенаправленного поведения через звено обратной афферентации осуществляется постоянная оценка реально полученного результата с тем, который был запрограммирован в акцепторе результата действия. Результат этой оценки и определяет дальнейшее поведение организма. Если реально полученный результат соответствует прогнозируемому, то организм переходит к следующему этапу деятельности. При их несоответствии в аппарате сличения возникает рассогласование, активирующее ориентировочно-исследовательскую деятельность организма.
Функциональная система условного рефлекса имеет два принципиально важных свойства: 1) полезный приспособительный результат, 2) логическая операциональная архитектоника, представленная конкретными механизмами с обязательной обратной афферентацией в центральную нервную систему о результатах действия.