

Тема 6. Активные электрические свойства биообЪеКтов
Цель: исследовать проведение возбуждения и параметры потенциала действия электровозбудимых тканей
Материалы и оборудование: осциллографы: С1-83, электростимулятор ЭСЛ-2, , шнуры питания, соединительные провода, ванночка с раздражающими электродами, соединительные провода, лягушка, чашка Петри, препаровальные инструменты, физиологический раствор, инструкции по эксплуатации приборов.
Вопросы для самоподготовки:
1) Механизмы формирования потенциала покоя и потенциала действия нервного волокна.
2) Роль потенциалозависимых ионных каналов в формировании мембранных потенциалов электровозбудимых клеток.
3) Методы регистрации мембранных потенциалов и ионных токов в электровозбудимых тканях.
Теоретическая часть
Раздражимость – свойство всех без исключения живых клеток, представляет собой способность реагировать на различные факторы внешней среды. Возбудимость – более узкое понятие и представляет собой способность реагировать на электрические стимулы, генерируя потенциал действия. К возбудимым тканям относят нервную, мышечную и железистую.
Потенциал покоя (ПП) большинства электровозбудимых клеток, и нервных в том числе, имеет калиевую природу, т. к. движение именно этих ионов по концентрационному и электрохимическому градиенту в основном обуславливает разность потенциалов на мембране в состоянии покоя.
ПП рассчитывается по формуле Гольдмана:
 (1)
(1)
где R универсальная газовая постоянная, T – абсолютная температура, F – число Фарадея, концентрация ионов К+, Na+, Cl- внутри [ ]i ,и вне [ ]o клетки, Р – проницаемость мембраны для ионов К+, Na+, Cl-.
При нанесении клетке электрического раздражения пороговой силы генерируется потенциал действия (ПД). При этом увеличивается проницаемость мембраны для ионов Na+, которые по концентрационному и электрохимическому градиенту поступают внутрь клетки. Таким образом, ПД имеет натриевую природу. Постоянство концентрации ионов К+ и Na+-внутри клетки обеспечивается механизмом активного транспорта. Его система представлена специальным ферментом-переносчиком Mg2+-зависимой Na+,К+- АТФ-азой. Этот переносчик осуществляет противоградиентный транспорт ионов К+ и Na+ через мембрану за счет энергии гидролиза АТФ.
Для изучения роли активного транспорта в электрических явлениях на мембране можно воздействовать либо непосредственно на переносчик, либо на метаболические процессы в клетке. Во втором случае изменяются процессы, связанные с синтезом АТФ, а это влечет за собой изменение в работе Na+,К+-АТФ-азы.
Для изучения роли ионов в развитии ПД самым простым способом является изменение электролитного состава физиологического раствора. Так, добавление ионов К+ согласно уравнению Гольдмана должно вызывать деполяризацию и сдвигать величину ПП в положительную сторону, изменение концентрации ионов Na+ в омывающем растворе также будет изменять параметры ПД.
Мембранную разность потенциалов нервных клеток можно регистрировать с помощью внутриклеточных и внеклеточных электродов. Объектами исследования в первом случае могут быть гигантские аксоны ряда беспозвоночных животных (например, кальмара), во втором случае нервные волокна позвоночных животных (в частности, лягушек).
При регистрации ПД нервного волокна лягушки с помощью внеклеточных электродов наблюдается двухфазный ПД. Монофазный ПД получается при блокировании проведения нервного импульса между отводящими электродами.
Нервные волокна, в том числе и седалищный нерв лягушки неоднородны по структуре. Они представляют собой набор нервных пучков, неодинаковых по радиусу, а, следовательно, и по скорости проведения возбуждения. В связи с этим, в нерве лягушки выделяют 3 группы волокон. Для волокон А-группы скорость проведения возбуждения составляет 27 м/с, для волокон В-группы 4,2 м/с, для волокон группы С 0,4 м/с. В свою очередь, волокна группы А делятся по скорости распространения возбуждения на 3 подгруппы: это , , -волокна. Скорость проведения возбуждения по этим волокнам составляет соответственно 42, 25 и 16 м/с. При внеклеточном отведении потенциалов нерва лягушки, изменяя напряжение на раздражающих электродах, можно наблюдать на экране осциллографа ПД , и -волокон. Сначала появляется ПД -волокон, затем и, наконец, всех трех видов (рис. 7).

Рисунок 7. Последовательность изменения амплитудно-фазо-частотных параметров ПД А-волокон седалищного нерва лягушки при увеличении напряжения на раздражающих электродах. А импульс артефакта; Т латентный период
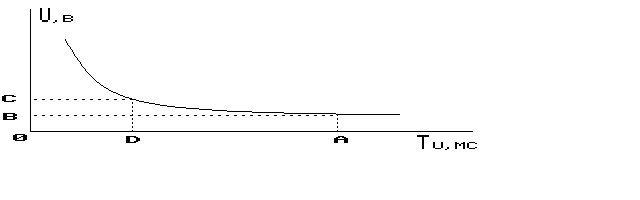
Рисунок 8. Кривая "сила длительность". Tu-длительность импульса в мсек, U (В) -амплитуда раздражающего импульса; ОД-хронаксия; ОВ-реобаза; ОА-полезное время.
Потенциал действия седалищного нерва лягушки при внеклеточном отведении подчиняется закону "все или ничего". При этом, если пауза между раздражающими импульсами больше рефрактерного периода, то, изменяя длительность импульса, можно получить кривую "сила длительность" и вычислить хронаксию и полезное время действия раздражающего тока (рис. 8).
Уменьшая длительность паузы между раздражающими импульсами, можно определить рефрактерный период для каждой из подгрупп А-волокон.
ПРАКТИЧЕСКАЯ ЧАСТЬ