

Множественный кроссинговер
В одной и той же тетраде может произойти два и более обмена между несестринскими хроматидами – двойной (множественный) кроссинговер.
При двойном – нужен анализ трех пар генов, гетерозиготных по двум аллелям.
Вероятность двойного кроссинговера (два независимых межгенных обмена) равна произведению вероятностей каждого из событий.
Если обнаружено 20% (р=0.2) кроссоверных гамет между генами А и В и 30% (р=0.3) между В и С, то вероятность двойного кроссинговера между генами А и В, В и С, приводящего к появлению кроссоверных гамет, равна 6% (0.2 х 0.3) Частота двойных кроссинговеров намного меньше одиночных.
Если три гена расположены довольно близко друг от друга, то ожидаемая частота двойных кроссинговеров чрезвычайно мала.
Если расстояние А В равно 3 ед, В С – 2 ед, то частота всего 0.06% (0.3 х 0.2).
Обнаружить это очень трудно, а при картировании четырех или пяти генов потомков нужно в разы больше
Генетическая карта – это последовательность маркеров в хромосоме и расстояния между ними, следующие из частот генетических рекомбинаций. Одна единица карты соответствует 1% рекомбинации или одному сантиморгану (1 сМ).
1)Генетическое расстояние между двумя маркерами (1сантиморган, сМ=1% рекомбинации).
Например, наблюдаемый процент рекомбинации в потомстве (12% рекомбинантов, как описывалось выше), где общее число
потомков равно 100%. Доля рекомбинантов по одной паре маркеров не может превышать 50% от общего числа потомков (несцепленное наследование);
2)Общая длина генетической карты организма (N cантиморган = N %) Рассчитывается как сумма минимальных экспериментально определенных генетических расстояний между парами маркеров, и для каждого вида своя.
Определение линейного расположения генов на хромосоме и расстояния между ними
Для проведения экспериментального скрещивания необходимы три условия:
1)Родитель, дающий кроссоверные гаметы, должен быть гетерозиготен по всем исследуемым локусам,
2)В потомстве должны точно определяться рекомбинантные фенотипы, поскольку генотипы гамет недоступны для прямого анализа. Каждый фенотип потомства должен соответствовать родительским либо рекомбинантным гаметам.
3)Для выявления всех кроссоверных фенотипов и картирования генов нужно проанализировать достаточное количество потомков
Исследуем три сцепленных с полом рецессивных мутантных гена y w eс.
Предположим такую последовательность их на Х- хромосоме и проверим это экспериментальным скрещиванием.
Самцы гемизиготны по всем 3 аллелям дикого типа, самки гомозиготны по всем 3 мутантным аллелям.
Фенотип самцов – дикий тип, в фенотипе самок – все 3 мутации.
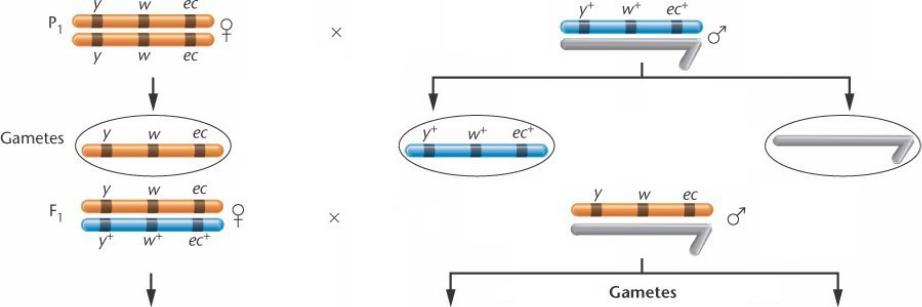
F1 – все самки – фенотип дикого типа (гетерозиготны), самцы –
мутантный фенотип (гемизиготны по мутантным аллелям, на У- хромосоме этих аллелей нет).
В результате кроссинговера самки F1 могут давать рекомбинантные гаметы.
Вданном скрещивании каждая из родительских гамет – либо все три аллеля, либо инертная У-хромосома.
ВF2 генотип этих гамет сразу же проявляется и у самцов, и
у самок, независимо от того, какие из гамет образуют зиготу.
Все 3 мутантных аллеля у исходных родителей на одной хромосоме, 3 нормальных – на гомологичной.
Возможны и другие варианты.
Если один из родителей несет аллели у и ес, а второй – w, то у гетерозиготных самок F1 мутантные аллели у и ес
будут на одной хромосоме, а w – на другой.
Второе условие соблюдено, поскольку самцы F1 образуют
гаметы либо с Х-хромосомой с 3 мутантными аллелями, либо с У-хромосомой без этих аллелей. Независимо от типов гамет самок F1 их генотипы будут проявляться у
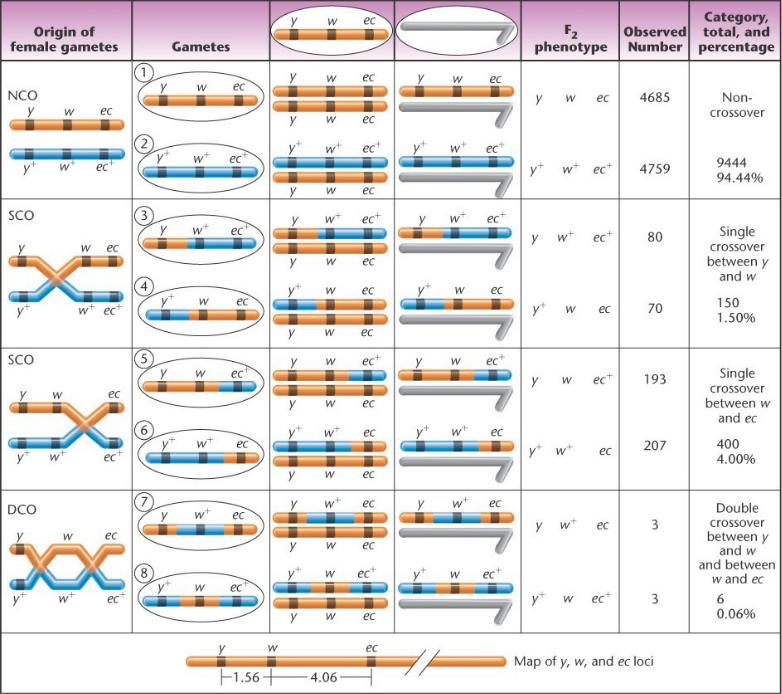
самцов F2 и у самок в потомстве этих самцов. Фенотипы мух F2 позволяют обнаружить кроссоверные и некроссоверные гаметы мух F1.
У самок F1 гаметы могут нести нерекомбинантные Х-хромосомы,
попадающие к потомству примерно в равных пропорциях. Эти гаметы – у гетерозиготных самок, поэтому в F2 будет два
комплементарных фенотипа – дикий и мутантный. Если одна Х-хромосома с одним мутантным аллелем, другая с двумя, то образуются реципрокные классы гамет и фенотипов.
Доля некроссоверных фенотипов в потомстве значительно выше, в данном опыте около 95%. Дважды кроссоверные гаметы
– в сумме только 0.06%.
Остальные 4 фенотипа – с одиночным кроссинговером.
Расстояние между локусами y w и w ec равно частоте всех рекомбинаций, обнаруженных между этими локусами, включая одиночные и двойные кроссоверы.
y w (1.5+0.06) =1.56 ед. карты , ec w (4.0+0.06) =4.06 ед. карты.