

Структура тРНК
Наиболее хорошо изучены небольшие стабильные тРНК. Всего 75-90 нуклеотидов, молекулярная структура у про- и эукариот почти одинакова.
4S тРНК образуется в результате процессинга из более длинных предшественников. У E coli тРНК, акцептирующая тирозин состоит из 77 нуклеотидов, ее предшественник – из 126.
1965 г. – Р. Холли расшифровал последовательность тРНК аланина. Оказалось, что ряд нуклеотидов уникален только для этой аланиновой тРНК. Одно из 4 оснований модифицировано в виде инозиновой кислоты, гипоксантина,
риботимидиновой кислоты и псевдоуридина. Эти модификации – необычные, редкие или непарные основания
появляются в ходе последующих посттранскрипционных изменений, а в процессе транскрипции в пре-тРНК встраиваются немодифицированные основания, которые изменяются под действием ферментов.

tRNA нуклеотиды могут содержать азотистые основания, модифицированные после
транскрипции.
Благодаря качанию, I (Inosine) и Im могут спариваться с U, C, или A.
Холли предложил двумерную модель молекулы тРНК в виде клеверного листа – несколько спаренных стеблей (за счет спаривания оснований) и неспаренных петель.
Поскольку GCU, GCC и GCA кодируют аланин, Холли попытался найти соответствующий антикодон в молекуле ала- тРНК и обнаружил его в виде CGI (3’-5’). Основание I (инозиновая кислота) может формировать водородные связи с U, С или А – третьими рибонуклеотидами в этих кодонах. В результате – антикодоновая петля
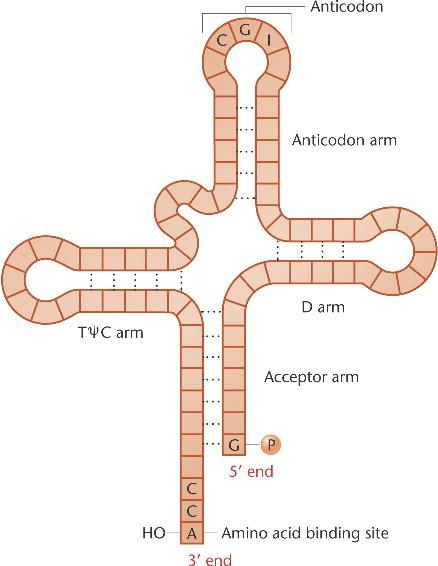
Молекула тРНК содержит много модифицированных оснований и содержит ряд стеблей и петель, сложенных в 3-мерную конфигурацию.
3‘-конец - сайт связывания аминокислоты, в то время как антикодоновая петля узнает кодон на мРНК.
Антикодон (CGI) этой молекулы tRNAala специфичен для
аминокислоты аланина, а также может ообразовывать пару с триплетами GCU, GCC, и GCA из- за качания.
Анализ структуры других тРНК выявил много общего.
На 3’ конце у всех тРНК есть последовательность ССА-3. К концевому остатку аденозина на этом конце ковалентно присоединяется аминокислота. На другом конце – 5’-G. Очень сходны размеры стеблей и петель. Каждая тРНК имеет антикодон, комплементарный соответствующему триплету, кодирующему определенную аминокислоту
Трехмерная модель тРНК – на основе кристаллографического исследования
Зарядка молекул тРНК
Для успешной трансляции молекулы тРНК должны присоединить соответствующие аминокислоты. Такая зарядка тРНК происходит с участием аминоацил-тРНК- синтетазы. Если разных аминокислот 20, то нужно не менее 20 разных тРНК и ферментов. Теоретически это число может соответствовать числу различных кодонов (61). Но качание третьего нуклеотида в кодоне снижает это количество до 32 тРНК и 20 синтетаз.
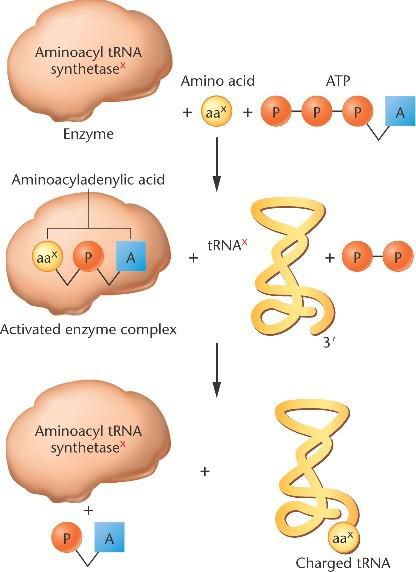
«Зарядка» тРНК начинается, когда аминоацил-тРНК-синтетаза катализирует превращение аминокислоты в аминоациладениловую кислоту.
Гидролизованная ATP отщепляет фосфат с образованием комплекса.
Аминокислота перемещается в соответствующую тРНК на 3‘-конец.
Такая заряженная молекула может участвовать в синтезе белка.
Аминоацил-тРНК-синтетазы – высокоспецифичные ферменты, узнающие только одну аминокислоту. Набор соответствующих аминокислоте тРНК называют изоакцепторными тРНК.
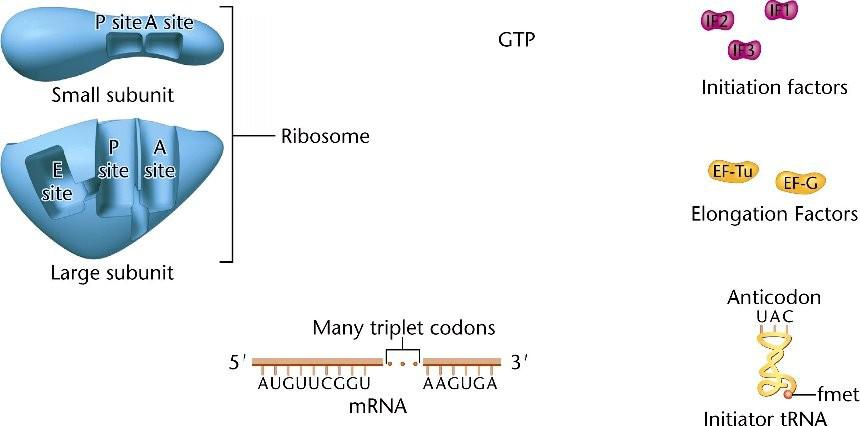
Инициация трансляции в E.coli затрагивает рибосомы, мРНК, энергоноситель GTP, несколько факторов инициации (IFs) и тРНК, которая несет антикодон UAC и заряжена модифицированной аминокислотой N-формилметионином (f-met).
E, P и А сайты на рибосоме.
Инициация трансляции.
Малая субъединица рибосомы связывается с несколькими факторами инициации (IF1, 2, 3); Этот комплекс связывается с мРНК.
Инициирующий tRNAfmet связывается с mRNA AUG кодоном в P (пептидильном) сайте. GTP гидролизуется, обеспечивая реакцию энергией.
Большая единица связывается с комплексом. Фактор элонгации Tu
связывается со следующей tRNA, содействуя вступлению в A (аминоацильный) сайт.