

Повторяющиеся кополимеры дают предсказуемые триплеты кодонов.
Эти синтетические мРНК могут быть использованы для встраивания
аминокислот в белки in vitro.
Почти универсальный генетический код служит словарем для перевода с языка мРНК на язык аминокислот.
Триплетный код обеспечивает 64 (43) кодона, чтобы определить 20 аминокислот.
Таким образом, код является вырожденным: многим аминокислотам соответствует более чем один кодон, только триптофан и метионин кодируются одним кодоном.
В дополнение к кодонам, которые определяют аминокислоты, есть один «старт» (или «инициирующий») кодон (AUG, который также кодирует метионин) и 3 "стоп" («терминирующих") кодона.
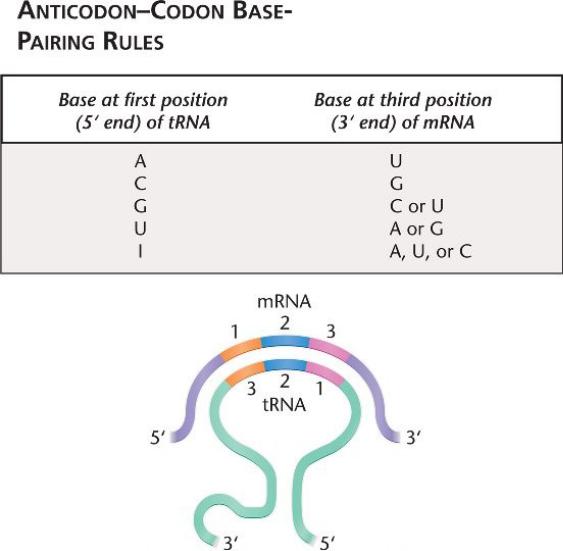
Во многих случаях первые две буквы генетического кода более критичны для определения аминокислоты.
Например, кодон валина (Val) зависит только от первых 2 букв (GU).
3-я позиция в кодоне может «качаться»: одна тРНК может
спариваться с более чем одним кодоном в мРНК.
U на 1-й позиции (5 ') антикодона тРНК может свариваться А или G на 3-й позиции (3') кодона мРНК, и G может также свапиваться с U или C.
Инозин (I), модифицрованное основание, обнаруженное в тРНК, может спариваться с C, U, или A.
Гипотеза качания предполагает, что для кодирования аминокислот 61 триплетом требуется не менее 30 различных тРНК.
Качание, как минимум, экономично не в ущерб точности, в бактериальных клетках 30-40 разных тРНК, в растительных и животных – до 50
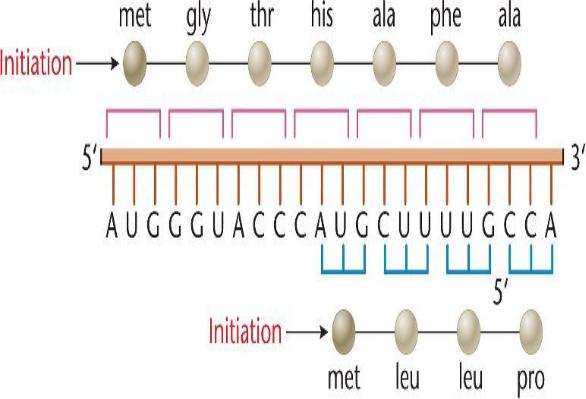
Инициация и терминация синтеза белка
Инициация синтеза белков высокоспецифична. Синтез всех
полипептидов начинается с модифицированного метионина – N- формилметионина (fmet).
Инициирующим кодоном служит только один триплет АУГ, кодирующий метионин. В других позициях иРНК этот кодон кодирует вставку обычного метионина. Изредка метионин в первой позиции кодируется триплетом ГУГ (в норме – валин) После завершения синтеза белка формильная группа или весь метионин отщепляются от полипептидной цепи. У эукариот первая аминокислота тоже метионин, но без формильной
группы Терминирующие кодоны УАГ, УАА и УГА. Они не узнаются
тРНК. Появление таких триплетов в кодирующей белок последовательности при мутации → прекращение синтеза белка → частичный синтез полипептида → преждевременный конец трансляции. Такие мутации в ДНК – нонсенс-мутации
Универсальность генетического кода
В1960-1978 гг. – считалось, что генетический код у всех организмов совершенно одинаков.
В1979 г. показано, что специфический код используется в митДНК человека и дрожжей. Затем исследовали и другие митДНК. Сравнение клонированных фрагментов митДНК с составом митохондриальных белков выявило несколько исключений из универсального генетического кода. Самое удивительное – терминальный кодон УГА в митДНК кодирует триптофан. Кодон АУА (в норме - изолейцин) в митРНК человека кодирует метионин, СУА в митРНК дрожжей треонин вместо лейцина
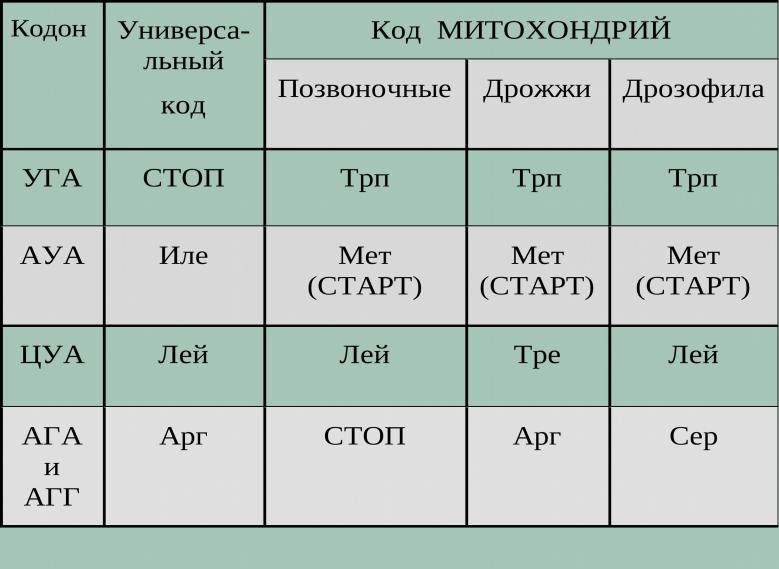
Отклонения от универсального кода в митохондриях разных организмов
1985 г. – у бактерий Mycoplasma и некоторых простейших жгутиковых – еще несколько исключений. Например, терминальный УГА – триптофан, или терминальный УАА и УАГ
– глутамин. Эти отклонения наиболее интересны, поскольку встречаются и у прокариот, и у эукариот
Изменение кодировки касается только третьей позиции – в цитоплазме АУА – изолейцин, в митохондриях – метионин, при этом метионин в цитоплазме АУГ. УГА в цитоплазме терминальный, в митохондриях – триптофан вместо обычного УГГ.
Предполагается, такие изменения появились в эволюции из-за уменьшения числа необходимых тРНК в митохондриях (всего 22 у человека)
Тем не менее кодовые различия – исключения из общих правил кодировки

Некоторые исключения из генетического кода находятся в митохондриальной ДНК (mtDNA), а также в ДНК некоторых одноклеточных организмов.
Некоторые изменения, такие, как кодон УГА в строке 1. включают только сдвиг в определении третьей, или «качающейся», позиции.