

Неклеточный синтез белков
Внеклеточной системе сборки аминокислот в полипептиды должны содержаться необходимые для синтеза белков факторы – рибосомы, тРНК, аминокислоты и другие компоненты, участвующие в трансляции + иРНК – матрица для трансляции
В1961г. иРНК еще не была выделена из клеток, но с помощью
полинуклеотид фосфорилазы синтезировали искусственную РНК, которую затем добавляли in vitro
Вотличие от РНК-полимеразы полинуклеотид фосфорилаза не нуждается в ДНК-матрице и добавляет рибонуклеотиды к растущей цепи случайным образом (соответственно их концентрации в смеси).
Вероятность встраивания определенного рибонуклеотида пропорциональна его доступности для фермента по сравнению с другими рибонуклеотидами
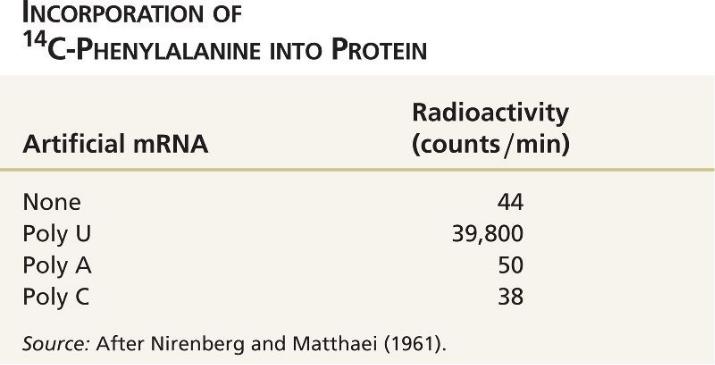
В качестве первого шага в расшифровке генетического кода были использованы короткие искусственные последовательности гомополимеров мРНК UUUUUU ..., AAAAAA ..., или CCCCCC в качестве матрицы для синтеза полипептидов с использованием радиоактивно меченых аминокислот.
Было обнаружено, что Poly U определяет включение 14С-фенилаланина, указывая, что кодон фенилаланина - UUU.
На матрице полиG прямого синтеза не получилось
Смесь кополимеров
Далее синтез гетерополимеров из двух и более рибонуклеотидфосфатов в разных соотношениях.
Предполагалось, что на основании относительных пропорций каждого рибонуклеозид фосфата можно прогнозировать частоту отдельных кодонов в синтетической РНК.
Если такую РНК затем ввести в систему, то можно определить долю каждой аминокислоты в синтезированном белке и состав триплетов, кодирующих специфичные аминокислоты
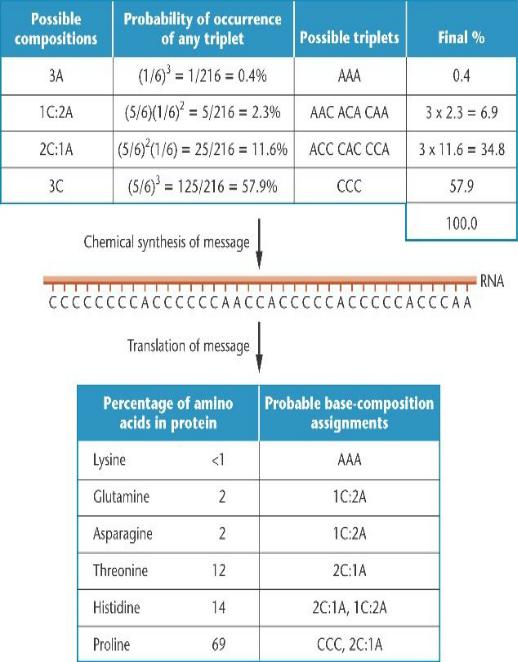
Расчет частоты возможных кодонов, полученных с использованием
гетерополимерной смеси в соотношении 1A: 5C.
Существует вероятность 1/6 для А и 5/6 - для C, чтобы занять каждую позицию в триплете.
Изучив процент аминокислот, включенных в синтезированный белок, можно предложить вероятный состав оснований для некоторых кодонов.
Пролин появляется в 69% случаев, поэтому он может кодироваться CCC
(57,9%) и одним кодоном, состоящем из 2C: 1A (11,6%).
Гистидин, (14%), вероятно, кодируется одним 2C: 1A (11,6%) и одним 1С: 2А (2,3%).
Треонин, (12%), скорее всего кодируется только одним 2C: 1А. Аспарагин и глутамин каждый, по- видимому, кодируются одним из 1С: 2A кодонов, и лизин - AAA.
Метод связывания триплетов
1964 г. – Ниренберг и Ледер предложили новый метод связывания триплетов, который помог установить точные последовательности кодонов
В присутствии молекул РНК рибосомы связываются с последовательностями РНК длиной 3 рибонуклеотида и формируют комплекс, напоминающий комплекс in vivo
Эти триплеты – кодоны иРНК комплементарны последовательностям тРНК, называемых антикодонами
Можно было получить заданную последовательность триплетов – специфическое связывание тРНК, несущей аминокислоту (аминоацил- тРНК) с триплетом РНК в комплексе иРНК-рибосома.
Аминокислоту метили – можно проследить какой из триплетов иРНК связывается с антикодоном
Инкубация меченой аминоацил-тРНК с синтезированными триплетами РНК и рибосомами на фильтре, задерживающем только крупные рибосомные комплексы.
При связывании меченой тРНК с триплетами иРНК на рибосомах метка оставалась на фильтре – соответствие тестируемой аминокислоты и данного триплета.
Так установили специфичные для каждой аминокислоты кодоны. В некоторых случаях добиться необходимого соответствия не удалось.
Было проверено 50 из 64 триплетов и получены два вывода
– одной аминокислоте соответствует несколько триплетов и один триплет кодирует только одну аминокислоту.
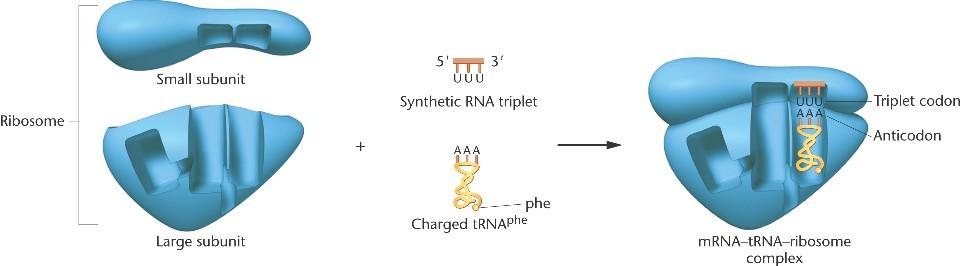
Анализ связывания триплетов.
Рибосомы могут связывать трехрибонуклеотидные последовательности (кодоны) in vitro, которые в свою очередь связываются с комплементарным антикодоном тРНК, переносящим специфичную радиоактивно меченную аминокислоту.
Весь комплекс может быть ассоциирован на нитроцеллюлозном фильтре и проанализирован по меченым аминокислотам.
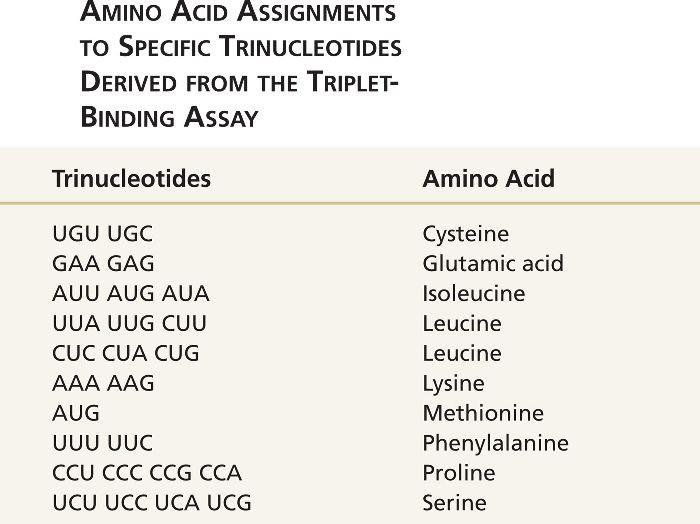
Около 50 из 64 кодонов были установлены с использованием анализа связывания триплетов.
Повторяющиеся кополимеры
Метод Кораны – синтез протяженных молекул РНК с заданной последовательностью, повторяемой несколько раз.
Например, последовательность из динуклеотидных повторов с двумя чередующимися триплетами – тринуклеотидные повторы образуют 3 потенциальных триплета, тетрануклеотидные – 4.
При добавлении таких синтетических иРНК в систему синтеза белков в синтезированных полипротеинах содержались теоретически ожидаемые пропорции аминокислот.

Корана (Nobel 1968) разработал методику для синтеза длинных молекул РНК, состоящих из коротких последовательностей (ди-, три-и тетрануклеотидов), повторяющихся много раз. Эти повторяющиеся кополимеры дают предсказуемые комбинации потенциальных кодонов.