

Несоответствие пар оснований происходит в одном из двух гомологов во время образования гетеродуплекса в мейозе.
Во время эксцизионной репарации, одно из двух несоответствий удаляется и синтезируется комплементарное основание, что приводит к возможной конверсии генов.
Если пара Г-С на одном из гомологов относится к мутантному аллелю, а пара А-Т к аллелю дикого типа. Конверсия пары Г-С в А-Т приведет к реверсии дикого типа
Генная конверсия позволяет объяснить явление у грибов – при скрещивании штаммов дикого типа и мутантных по этому же гену штаммов образуются равные количества нормальных и мутантных спор, но иногда бывает 3 к 1 или 1 к 3. Генная конверсия встречается также в процессе митоза у грибов и дрозофилы
Генетический код
Центральная догма
репликация
транскрипция трансляция
 ДНК
ДНК РНК
РНК белок
белок
Передача |
Реализация информации |
информации |
в одной клетке |
в поколениях |
|
делящихся |
|
клеток |
|
Центральная догма
репликация
транскрипция трансляция
 ДНК
ДНК РНК
РНК белок
белок
Принцип |
Компле- |
Генетический |
копирования |
ментарность |
код |
Генетический код
•Способ записи информации о первичной структуре белков через последовательность нуклеотидов ДНК и РНК.
•«Словарь» перевода с языка нуклеиновых кислот на язык белков.
•Полностью расшифрован к 1966
Первые представления о генетическом коде
Еще в 50-х гг. считалось, что ДНК кодирует белки в процессе своего синтеза без посредников.
Предполагалось, что информация из ДНК в ядре переносится к рибосомам, РНК которых и служит матрицей для синтеза белков, однако было показано, что имеется нестабильная молекула-посредник, а рРНК - стабильная молекула.
В 1961 г. Ф. Жакоб и Ж. Моно доказали существование матричной РНК или РНК-посредника. После открытия мРНК стало ясно, что генетический код из ДНК транслируется в аминокислотную последовательность посредством РНК.
Обоснование триплетности кода Гамовым
Моноплетный
Дуплетный
Триплетный
н. а.к. |
Сколько а.к. можно |
|
закодировать |
1 → 1 |
4 |
2 → 1 |
16 |
3 → 1 |
64 |
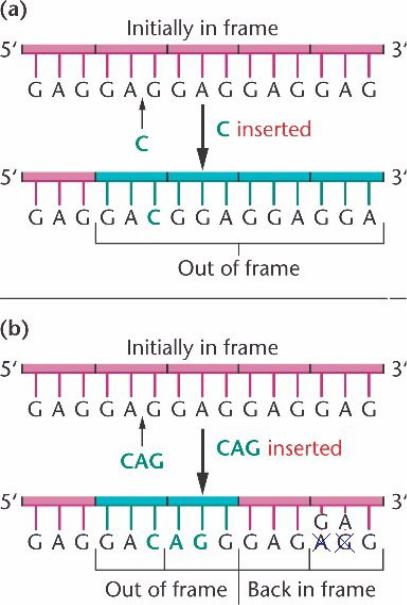
Мутация одиночной нуклеотидной вставки в ген фага T4 вызывает сдвиг рамки считывания всех последующих кодонов, и потенциально неправильные аминокислоты.
В результате белки, как правило, нефункциональны, и фаг T4 с такой мутацией рамки считывания не может репродуцироваться в E. coli.
Если три вставки происходят вместе, рамка считывания восстанавливается после вставки.
Такая мутация производит белок с достаточной функциональностью, чтобы
восстановить способность фага к заражению E. coli.
Триплетность кода подтверждена экспериментально – делеция или вставка трех нуклеотидов не сдвигает рамку считывания последовательности ДНК во время транкрипции у фага T4 , если нуклеотидов один или два, то возникает мутация
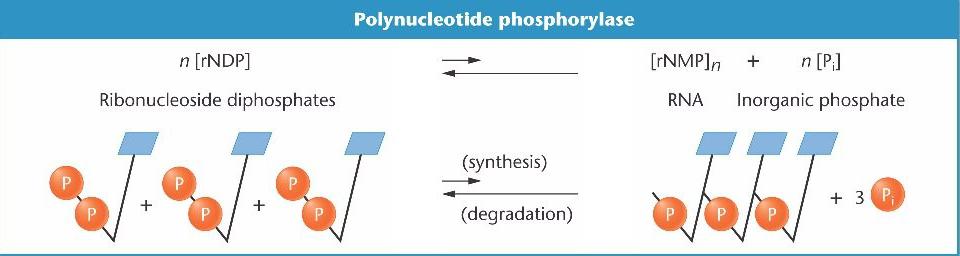
Расшифровка кода
Ниренберг и Маттеи (Nobel 1968) производили последовательности РНК с высокой концентрацией рибонуклеозид дифосфатов. В эксперименте использовалась неклеточная система синтеза белков in vitro вместе с ферментом полинуклеотид фосфорилазой, участвующей в синтезе иРНК, которая стала матрицей для синтеза белков
Продуцированную последовательность нуклеотидов можно регулировать путем изменения концентрации rNDPs, и она служит как бы "посредником« для синтеза полипептидных цепей in vitro..