

В дальнейшем показано, что теломераза добавляет TTGGGG даже к концам хромосом, не несущим эту последовательность, т.е. эта последовательность не служит сигналом для активации теломеразы. Теломераза содержит в своей молекуле сегмент РНК, образующий рибонуклеопротеин, важный для каталитической активности. Этот фрагмент РНК кодирует последовательности используемые ферментом как матрица. Он содержит 159 нуклеотидов, включая 5’-ААСССС-3’, комплементарный повтору в шпилечной структуре. РНК-содержащая теломераза напоминает обратную транскриптазу, поскольку синтезирует на РНК-матрице комплементарную последовательность ДНК
При старении клеток происходит укорачивание теломер. В 1961 г. Л. Хейфлик обнаружил, что соматические клетки могут делиться только ограниченное число раз. Хейфлик показал, что фибробласты клеток кожи делятся примерно 50±10 раз, после чего останавливаются. В большинстве соматических клеток эукариот теломераза неактивна, и поэтому с каждым клеточным делением теломеры становятся все короче. После многих делений они укорачиваются настолько, что клетка теряет способность к делению. Опухолевые клетки сохраняют теломеразную активность и делятся без ограничений.
Теломераза
•фермент, надстраивающий концы хромосом, содержит РНК.
•удлинение происходит путем
обратной транскрипции:
РНК → ДНК
На концах хромосом находятся длинные некодирующие повторы 5’ – ГГТ ТАГ – 3’
10-15 тысяч н.п. у человека
•Теломераза активна в клетках
–зародышевого пути
–эмбриональных
–стволовых
–раковых – поэтому они бессмертны
•Теломераза неактивна
–в соматических клетках – ген для нее
там, конечно же, есть, но выключен
Рекомбинация ДНК
Генетические изменения в одинаковых участках двух гомологичных хромосом относятся к так называемой общей или гомологичной рекомбинации. Предложено несколько моделей кроссинговера, имеющих много общего
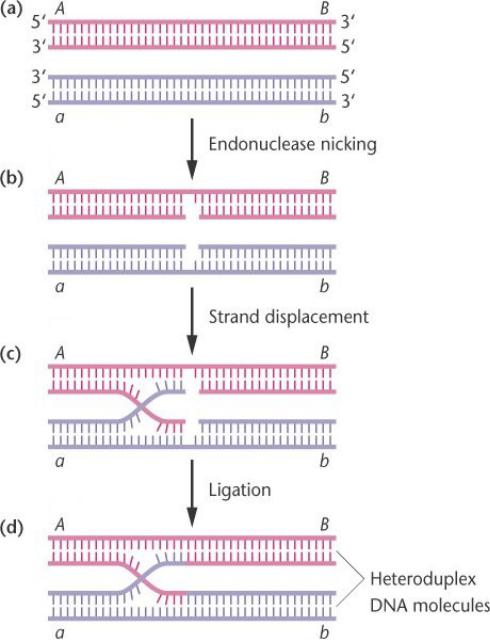
Гомологичная рекомбинация в течение мейоза.
Два дуплекса ДНК, которые имеют гомологичные последовательности, спариваются.
Эндонуклеазные надрезы производят одноцепочечные бреши на одинаковых позициях. Одиночные цепи смещаются и спариваются с комплементарными участками на противоположном дуплексе. Лигаза заделывает свободные концы, создавая гетеродуплексные молекул ДНК.
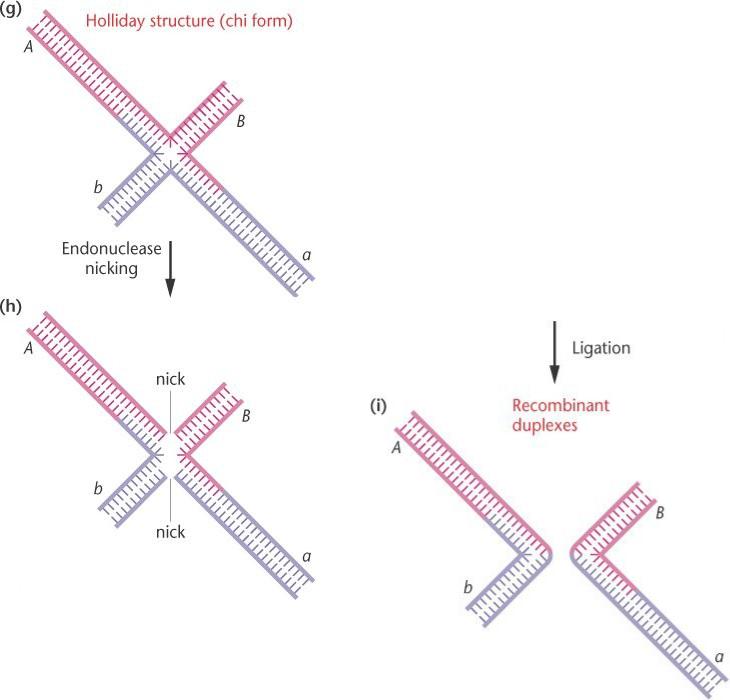
Гомологичная рекомбинация.
Нижние части поворачиваются на 180°, что создает плоскую χ (chi) форму (структуру Холидея).
Два другие цепи в это время надрезаны эндонуклеазой.
Образуется крестообразная структура Холидея
Бреши
закрываются лигазой, образуя
рекомбинантные дуплексы.
После разделения дуплексов и поворота их по отношению друг к
другу на 180о образуется промежуточная плоская структура, называемая хи-формой Если две не вовлеченные в обмен цепи
гомологов разрезаются эндонуклеазой и лигируются, то возникают рекомбинантные дуплексы. В результате на хромосомах изменяется последовательность аллелей
Генная конверсия
Генная конверсия происходит между двумя тесно сцепленными генами, и она нереципрокна При скрещивании двух штаммов нейроспоры, несущих разные мутации (а+ х b+) между этими генами может произойти реципрокная рекомбинация, тогда образуются споры с генотипами ++ и аb. При нереципрокной рекомбинации в потомстве от такого скрещивания будет генотип ++, а генотип ab отсутствует. Это происходит за счет генной конверсии, частота которой выше частоты мутаций в данных генах. Считается, что один из аллелей как бы конвертирует (превращается) в другой аллель путем генетической рекомбинации.
Генная конверсия – следствие рекомбинации, в процессе которой основания спариваются ошибочно. Неправильно спаренные отрезки гетеродуплекса могут репарироваться путем вырезания и синтеза комплементарной последовательности
Возможны два способа коррекции – ошибочно спаренные нуклеотиды вырезаются, и восстанавливается исходная последовательность или же коррекция происходит путем синтеза нуклеотидов, комплементарных измененным основаниям, что происходит к нуклеотидным заменам.
В результате конверсии вместо различных аллелей, локализованных на двух гомологичных хромосомах, появляются идентичные аллели