

ДНК-полимераза исправляет ошибки
Если новый нуклеотид не спарен – фермент не может двигаться дальше.
Тогда он выедает неверный нуклеотид и ставит другой.
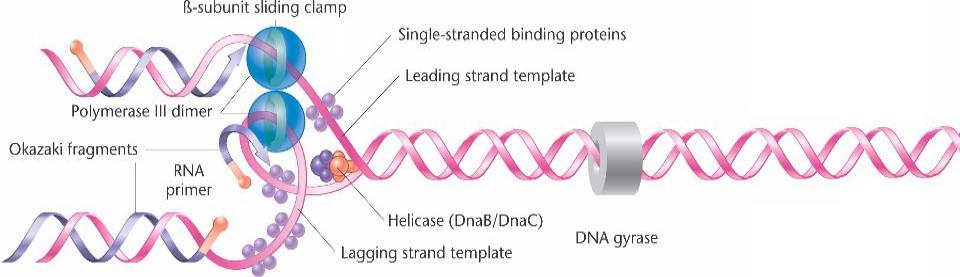
Основные этапы синтеза ДНК в бактериях.
При продвижении вилки белки геликазы раскручивают двойную спираль. Праймаза устанавливает короткий РНК праймер.
Каждый коровойфермент димера ДНК-полимеразы III связан с матричной цепью бета-субъединицей по типу скользящего замка.
Синтез непрерывен на лидирующей цепи, а на отстающей цепи происходит в виде фрагментов Оказаки.
Отстающая цепь образует петлю для возможности одновременного синтеза обеих цепей.
ДНК-полимераза I заменяет РНК праймер (синтезированный праймазой) на ДНК.
ДНК-лигаза сшивает фрагменты Оказаки.
Синтез ДНК у эукариот
Все то же самое, но сложнее из-за огромного количества ДНК у эукариот
Множественные точки начала репликации
Репликация у эукариот начинается во множестве точек. Если у дрожжей геном содержит 250-400 репликонов, то у
млекопитающих не менее 25000. Выделенные из дрожжей точки начала репликации назвали автономно реплицирующимися последовательностями (ARS – autonomously replicating
sequences).
Синтез ДНК у эукариот проходит в S-фазу клеточного цикла, в течение этой фазы происходит последовательная активация кластеров из 20-80 соседних репликонов, пока не будет реплицирована вся ДНК.
Эукариотические ДНК-полимеразы
Выделено 6 форм полимераз.
Особенно важны 3 – Pol , , , две другие - , , по-видимому, вовлечены в репарацию ДНК. Шестая - участвует в синтезе митДНК, хотя и кодируется ядерным геном. Все, кроме , состоят из множества субъединиц, выполняющих в процессе репликации различные функции
альфа, бета, гамма, дельта, эпсилон, зита
Полимеразы и особенно важны для синтеза ядерной ДНК. Две из 4 субъединиц -полимеразы служат праймазами для синтеза РНК-затравок на ведущей и отстающей цепях ДНК Этот фермент обладает низкой процессивностью, т.е. синтезирует короткую последовательность, а затем диссоциирует К РНК-праймеру добавляется затем короткая последовательность ДНК и происходит переключение полимеразы.
Pol диссоциирует, освождая место для . Этот фермент обладает высокой процессивностью и экзонуклеазной активностью 3’-5’, т.е. способен к проверке ошибок репликации. Скорость синтеза второго фермента в 100 раз выше, чем у первого. Pol участвует в элонгации и проверке ошибок на растущей цепи ДНК.
Множество репликонов у эукариот – больше молекул ДНК- полимераз. У E coli 15 молекул ДНК-полимеразы III, в животной
– до 50000 копий полимеразы . Низкая скорость репликации у эукариот компенсируется множеством коротких репликонов. Если геном E coli реплицируется за 20-40 мин, то гораздо больший дрозофилы – за 3 мин
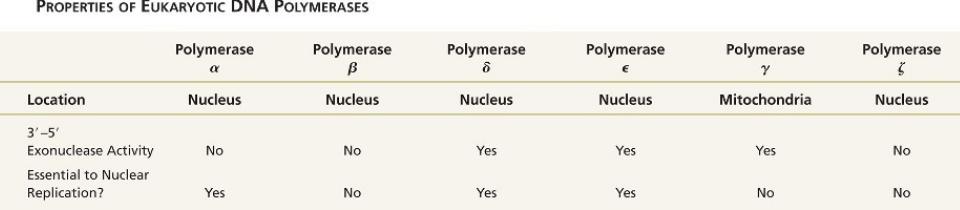
Три эукариотических ДНК полимеразы катализируют реакцию репликации ДНК,
остальные вовлечены в репарацию.
Pol α (alpha) синтезирует РНК праймеры в течение инициации. Потом, в процессе, называемом переключением полимеразы, она заменяется Pol δ (delta), которая выполняет основную задачу одновременного удлинения обеих цепей.
Pol ε (epsilon) является другим ферментом, участвующим в ядерном синтезе ДНК, возможно, играет роль в связывании с точкой origin или синтезе отстающей цепи.
Pol γ (gamma) кодируется ядерным геном, хотя его функция заключается в синтезе митохондриальной ДНК.
Репликация ДНК, теломеры и теломераза
В отличие от кольцевых бактериальных хромосом у эукариот они линейны, концы называются теломерами. Когда процесс
синтеза ДНК подходит к концу ведущей цепи, на отстающей цепи после удаления РНК-праймера возникают трудности. Появившаяся брешь заполняется нуклеотидами, присоединяющимися к 3’-ОН группе в процессе прерывистого синтеза. На конце хромосомы нет гидроксильных групп, поэтому в ходе каждого этапа синтеза ДНК теоретически должно происходить укорочение хромосомы на длину праймера. Механизм решения этой проблемы подсказало открытие теломеразы.
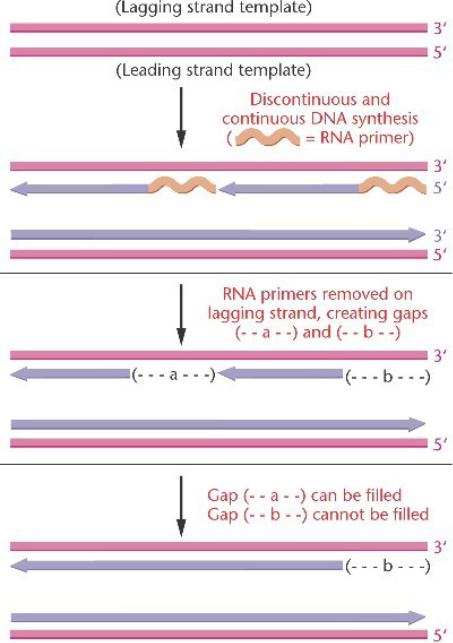
Полуконсервативный синтез лидирующей цепи в линейной
хромосоме может происходить нормально до конца двойной спирали.
На отстающей цепи, после удаления последнего РНК праймера, нет свободной 3'-OH
группы для ДНК-полимеразы, чтобы продолжать элонгацию.
Брешь на отстающей цепи приводит к укорочению хромосомы после каждого раунда синтеза.
Это укорочение хромосомы может играть роль старения в соматических клетках, и его следует избегать в половых клетках.
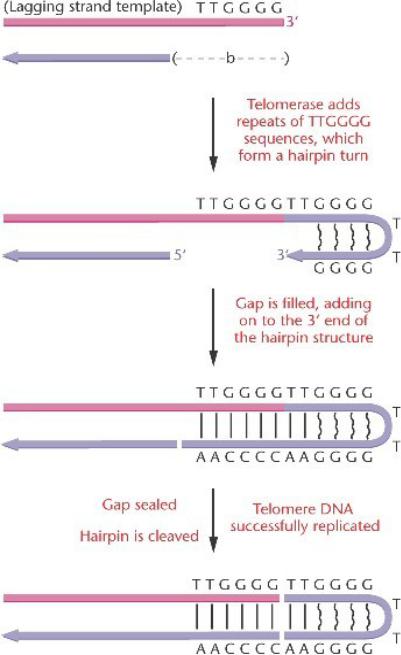
Теломерная ДНК на концах линейных хромосом эукариот состоит из коротких тандемных повторов: последовательность TTGGGG повторяется много раз.
Они синтезируются с помощью фермента теломеразы.
Фермент теломераза может синтезировать короткие последовательности ДНК (теломеры) на 3‘-конце эукариотических хромосом, предотвращая укорочение хромосом в половых клетках.
Этот фермент добавляет повторы TTGGGG последовательности, которые загибаются на себя, образуя необычные водородные связи G-G.
Брешь заполняется ДНК-полимеразой и лигазой.
Петля шпильки, затем отщепляется, сохраняя первоначальный дуплекс.
Это позволяет гаметам и злокачественным клеткам, а также некоторым "бессмертным" культурам клеток продолжать дубликацию линейной ДНК.