

Модель синтеза ДНК включает несколько этапов:
Раскручивание спирали ДНК
У E. coli участок начала репликации состоит из 245 п.н. с повторами из 9 и 13 оснований, называемых 9- и 13-мерами. Инициируется раскручивание белком DnaA. Этот белок связывается к каждым из 9- меров, затем с ДНК связываются белки DnaB и DnaC, что приводит к дальнейшему раскрытию спирали. Открытая конформация ДНК стабилизируется с помощью белков, связывающихся с односпиральной ДНК – SSBP- белков (single-stranded binding proteins).
По мере раскручивания в нераскрученной части возникает суперскрученность, которая ослабляется ДНК-гиразой – ферментом группы ДНК-топоизомераз, которая, разрезая цепи ДНК, удаляет лишние обороты и узлы.
Молекула ДНК, полимеразный комплекс и ферменты репликации образуют комплекс, называемый реплисомой.
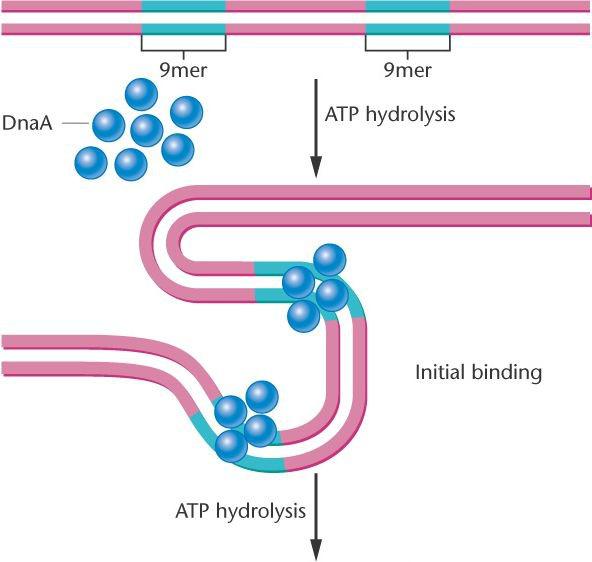
Раскручивание
бактериальной спирали начинается, когда
мономеры белка геликазы DnaA
связываются с сайтами ДНК, содержащими повторяющиеся последовательности из
9 и 13 оснований (9mers и 13mers).
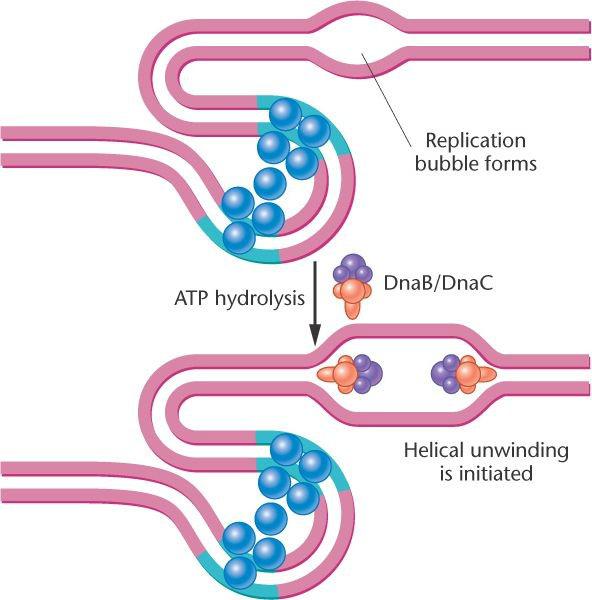
Белки геликазы DnaB и
DnaC открывают спираль разрывая водородные
связи между основаниями, денатурируя двойную
спираль и образуя
репликационный пузырь.
Энергия для разрыва
водородных связей
обеспечивается гидролизом
ATP.
Инициация синтеза ДНК
После раскручивания небольшого участка ДНК начинается синтез новой цепи. Для элонгации цепи ДНК-полимеразе III требуется праймер со свободным 3’-концом. Хотя у РНК нет свободной 3’-гидроксильной группы, праймером для инициации синтеза ДНК служит РНК.
Сначала на ДНК-матрице с помощью РНК-полимеразы (праймазы) синтезируется короткий (5-15 нуклеотидов) комплементарный фрагмент РНК. Для этого не требуется свободный 3’-конец. К этому фрагменту РНК ДНК-полимераза III добавляет 5’-дезоксирибонуклеотиды. ДНК-полимераза I вырезает праймер, замещая его ДНК.
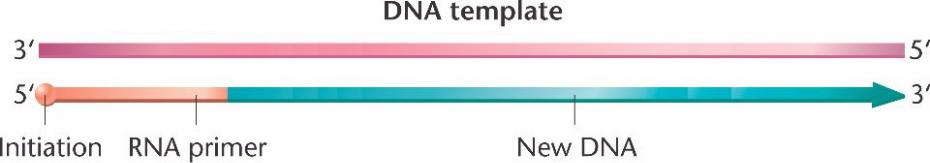
Инициация синтеза ДНК начинается, когда праймаза строит короткий РНК праймер в направлении 5' к 3', комплементарно матричной цепи спирали.
Прерывистый и непрерывный синтез ДНК
Известно, что цепи в ДНК антипараллельны – одна от 3’ к 5’- концу, другая от 5’ к 3’. ДНК-полимераза III осуществляет только синтез 5’-3’, поэтому синтез в пределах репликационной вилки происходит одновременно в двух направлениях, спираль расплетается и вилка продвигается. Матрицей для непрерывного синтеза служит только одна цепь – ведущая или лидирующая. С продвижением репликационной вилки на другой или отстающей цепи образуются все новые и новые точки инициации прерывистого синтеза ДНК.
Образуются фрагменты длиной 1000-2000 нуклеотидов с РНК- праймером – фрагменты Оказаки, которые с помощью ферментов
сшиваются с удалением РНК-праймеров. Удаление праймеров и синтез недостающих нуклеотидов проводит ДНК-полимераза I, сшивку фрагментов – ДНК-лигаза.
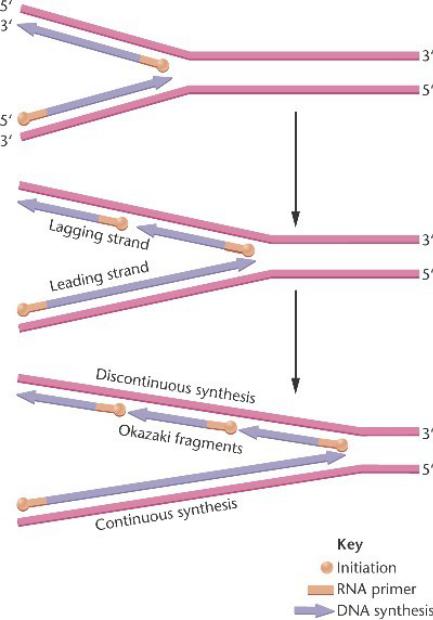
Поскольку полимеризация с участием ДНК полимеразы III происходит только в
направлении от 5' к 3', элонгация вдоль двух
антипараллельных цепей становится различной.
Синтез вдоль лидирующей цепи
репликационной вилки может происходить непрерывно.
Синтез вдоль отстающей цепи должен быть прерывистым, в виде фрагментов Оказаки,
каждый с праймером РНК.
Конкурентный синтез ведущей и отстающей цепей ДНК
Репликация ДНК происходит одновременно на двух цепях. Отстающая цепь образует петлю, изменяя направление синтеза физически, но не биохимически. Фермент-димер проводит репликацию на обеих цепях. После завершения синтеза фрагмента петли происходит образование новой петли, и процесс синтеза очередного фрагмента продолжается. Кроме того, в состав фермента входят субъединицы, не позволяющие основному
ферменту сползать с матрицы в процессе репликации, –
скользящий замок.
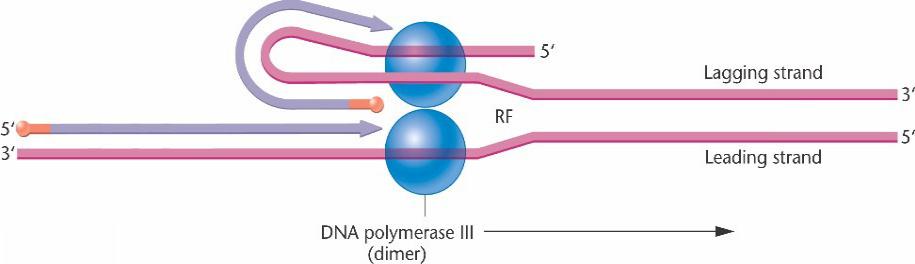
Полимеризация происходит одновременно на обеих цепях одним холоферментом ДНК-полимеразы III. Отстающая матричная цепь делает петлю на репликационной вилке, что позволяет каждому коровому ферменту димера добавлять основания в направлении от 5 'к 3'.
β (beta) субъединица формирует димер, который служит "замком«, сохраняющим связь корового фермента с матрицей ДНК.
Таким образом, весь холофермент движется вдоль родительского дуплекса как скользящий замок, продвигая репликационную вилку.
Проверка и коррекция ошибок в процессе репликации ДНК
Возможные ошибки репликации узнаются и вырезаются ДНК- полимеразами II и III с 3’-5’-экзонуклеазной активностью. Экзонуклеазная коррекция повышает точность синтеза ДНК