

Тест Эймса подвергает ауксотрофные штаммы сальмонеллы (his–)
воздействию химических соединений, чтобы оценить их мутагенность.
Экстракт печени добавляют, так как метаболические изменения этих соединений в печени может сделать их мутагенными.
Мутагенность химических веществ находит свое отражение в частоте обратных мутаций, которые дают прототрофы дикого типа (his+).
Исправление повреждений ДНК: системы репарации
Фотореактивная репарация у прокариот
При исследовании мутагенности УФ было обнаружено множество типов репарации повреждений ДНК.
В 1949 г. А. Кельнер открыл репарацию индуцированных ультрафиолетом нуклеотидных сшивок – фотореактивацию.
Повреждения ДНК, индуцированные у E. coli УФ, можно частично восстановить, если после облучения поместить бактерии под синий свет. Фотореактивная реакция зависит от температуры, т.е. это ферментативная реакция.
Позднее показано, что репарация зависит от активности фермента –
фермента фотореактивации (PRE). Этот фермент расщепляет ковалентные связи между остатками Т в тиминовых димерах и восстанавливает структуру ДНК. Для активации молекулы фермента требуется фотон, но затем свет не нужен. Клетки E. coli могут обойтись без этого фермента, поскольку нуль-мутация в гене, кодирующем этот фермент, не летальна.
У человека и других эукариот такой системы фотореактивации нет.

Фотореактивная
репарация.
В E. coli фермент фотореактивации (PRE) может расщеплять связи между тиминовыми димерами, заряжаясь от фотона синего света.
Реакция обратна действию УФ-излучения на ДНК.
Эксцизионная репарация у прокариот и эукариот
В 60-е годы у прокариот обнаружена и другая зависимая от света
система репарации повреждений ДНК. В основе ее лежит механизм эксцизионной репарации, который у прокариот и эукариот включает
три основных этапа:
1)нуклеаза узнает и вырезает ошибочное основание или измененную структуру ДНК. Такая эксцизия затрагивает одно основание или нуклеотид или несколько нуклеотидов, смежных с ошибочным. В спирали ДНК остается брешь.
2)ДНК-полимераза I заполняет эту брешь дезоксирибонуклеотидами, комплементарными нуклеотидам в неповрежденной цепи ДНК, которые пристраиваются с 3’-ОН конца.
3)ДНК-лигаза сшивает последний разрез цепи, который остается на 3’-ОН конце последней пары оснований, и закрывает брешь.
Известно два типа эксцизионной репарации – репарация оснований и репарация нуклеотидов.
При эксцизионной репарации оснований исправляются повреждения азотистых оснований, вызванные спонтанным гидролизом или химическими агентами. Первый этап такой репарации в клетках E. coli включает узнавание химически модифицированные основания ДНК- гликозилазами, специфичными для разных повреждений ДНК. Эти ферменты расщепляют связь между основанием и сахаром, и появляется апиримидиновый сайт (АР-сайт), который узнается АР- эндонуклеазой. Эндонуклеаза разрезает цепь ДНК в АР-сайте, а разрез узнается ферментами эксцизионной репарации, исправляющим это повреждение.
С помощью эксцизионной репарации нуклеотидов исправляются блоки ошибок, изменяющие структуру двойной спирали, например пиримидиновые сшивки.
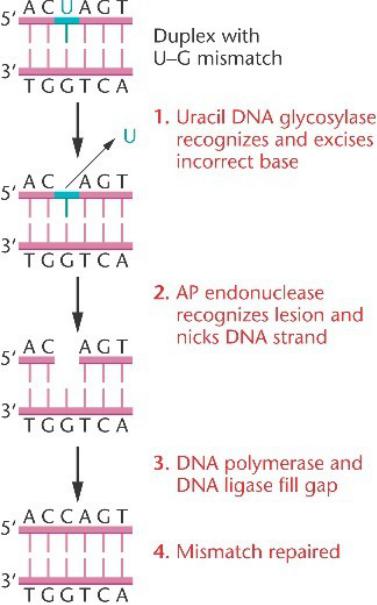
Репарация оснований в E.coli.
Несоответствие пар оснований узнается ДНК гликозилазой, которая отделяет основание от сахара, создавая апуриновый / апиримидиновый (AP) сайт.
AP эндонуклеаза делает разрез в фосфодиэфирном остове на месте AP сайта.
Этот разрыв заполняется ДНК-полимеразой I и ДНК-лигазой
Ошибка исправлена.
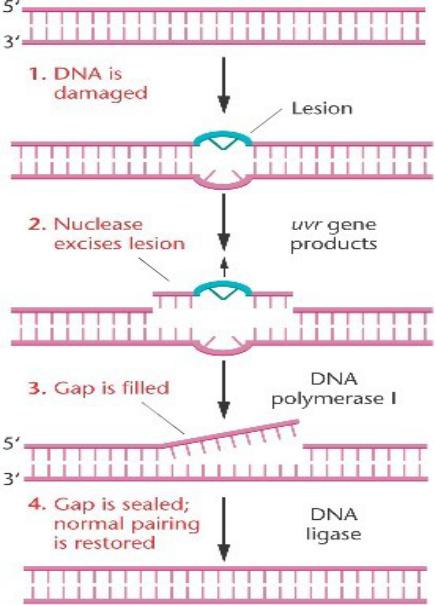
Эксцизионная репарация нуклеотидов.
Поврежденная ДНК узнается УФР (УФ репарационными) белками.
Несколько нуклеотидов вырезается вокруг всего поражения нуклеазой.
Этот разрыв заполняется ДНК- полимеразой I и ДНК-лигазой.
Ошибка исправлена.
Репарация ошибок репликации
Для исправления ошибок репликации ДНК существуют другие системы репарации. Функцию коррекции таких ошибок выполняет ДНК-полимераза III. В процессе полимеризации ДНК в молекулу могут встраиваться некомплементарные нуклеотиды, которые распознают и вырезают из цепи ДНК ферментным комплексом, а затем неправильный нуклеотид замещается комплементарным. У бактерий коррекция ошибок повышает точность синтеза ДНК на два порядка – первоначально некомплементарные нуклеотиды встречаются с частотой 10-5, после коррекции – с частотой не более 10-7.
Для исправления ошибок оставшихся после коррекции существует дополнительный механизм репарации ошибок спаривания. При повреждениях ДНК или неправильном спаривании оснований эти ошибки должны узнаваться, а неправильный нуклеотид или нуклеотиды удаляться и замещаться комплементарными.