

Молекулярная основа мутаций
Любое нарушение нуклеотидной последовательности – мутация.
Замещение пуринового основания пуриновым, пиримидиновое – пиримидиновым – транзиция, замена пурина на пиримидин и наоборот
– трансверсия.
Возможна инсерция (встраивание дополнительного основания) или делеция, что приводит к изменению кода и сдвигу рамки считывания

Точечные мутации являются заменами оснований, в которых одна пара нуклеотидов изменяется.
Вставки и делеции могут привести к мутациям сдвига рамки считывания, где все последующие аминокислоты изменяются.
Мутация сдвига рамки, кратная трем нуклеотидам, может восстановить начальные рамки считывания.
Таутомерные сдвиги
В 1953 г. Уотсон и Крик обнаружили, что основания в составе ДНК имеют альтернативную химическую структуру или таутомерную форму.
Предположили, что мутации в паре оснований могут быть обусловлены таутомерными сдвигами. Наиболее стабильные таутомерные формы образуют обычные водородные связи, которые удерживают две спирали ДНК. Реже встречаются таутомеры, образующие водородные связи с некомплементарными основаниями, хотя такое спаривание и происходит между пурином и пиримидином.
Во время репликации ДНК редкие таутомерные формы оснований спариваются с некомплементарными основаниями, а в процессе следующей репликации каждое из этих неправильно спаренных оснований связывается с комплементарным пурином или пиримидином, в результате – транзиция.

Нуклеотиды могут существовать в таутомерных формах (структурных изомеров) из-за изменения положения протона (таутомерного сдвига). Реже, переходные таутомеры
могут образовывать водородные связи с некомплементарными основаниями. Аномальное спаривание всегда происходит между пиримидином и пурином, как показано в T-G и C-A-парах.

Мутация транзиции.
Редкие таутомеры в матричной цепи спариваются с некомплементарными основаниями во время репликации ДНК.
В этом примере, аномальная T - A пара образует аномальную C - A пару в одной дочерней спирали.
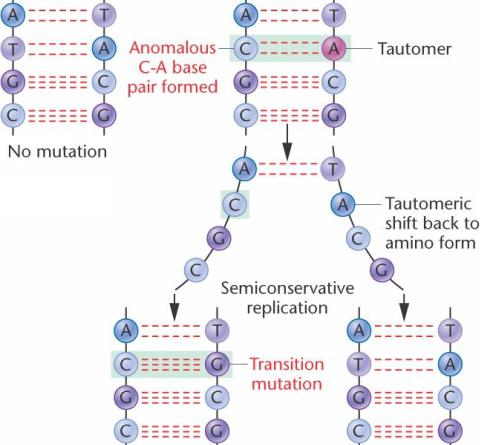
В следующем раунде репликации, несоответствующие члены пар оснований разделяются; таутомер обычно смещается назад к своему нормальному изомеру.
Репликация 2 цепей при обычном спаривании оснований приводит к
точечной мутации, называемой
мутацией транзиции, при которой пурины заменяются пуринами, или пиримидины заменяются на пиримидины.
В этом примере, T - A пара дикого типа мутировала в C - G.
Аналоги оснований
В процессе репликации ДНК обычные пиримидиновые и пуриновые основания могут замещаться химически модифицированными аналогами. Пример – производное урацила 5-бромурацил, соединяющийся с дезоксирибозой, образуя бромдезоксиуридин. Наличие брома вместо метильной группы увеличивает вероятность таутомерного сдвига и транзиции пары А-Т в пару Г-Ц. Есть и другие – 2-аминопурин – аналог аденина, который спаривается с Т или Ц и приводит к аналогичной транзиции.
Поскольку такие аналоги индуцируют мутации типа транзиций, их используют для индукции обратных мутаций, т е реверсии к дикому типу.
Реверсии протекают и спонтанно, но с гораздо меньшей частотой

5-бромурацил (5-BU) - производное урацила и ведет себя как аналог тимина, который увеличивает вероятность таутомерного сдвига от нормальной кето формы в енольную форму, неправильно спариваясь с гуанином вместо аденина.
После одного раунда репликации результат мутации транзиции - A - T в G - C.

Алкилирующие соединения
Один из примеров – горчичные газы – серосодержащие мутагены, известные с Первой мировой войны.
Механизм – метильная или этильная группа, присоединяющаяся к амино- или кетогруппе нуклеотидов. Сейчас основной агент – этилметансульфонат, алкилирующий кетогруппы Г и Т.
Мутация также типа транзиции.
Акридиновые красители и мутации сдвига рамки
Другие химические мутагены вызывают мутации сдвига рамки считывания, обусловленные вставкой или делецией одного или нескольких нуклеотидов. Индукция таких мутаций – на примере акридиновых красителей – профлавина и акридинового оранжевого, молекулы которых близки по размерам к нуклеотидам и могут встраиваться между пуринами или пиримидинами в молекуле ДНК. Это приводит к искривлению двойной спирали и к появлению делеций или инсерций.
Предполагается, что мутации сдвига рамки появляются вследствие брешей, возникающих в молекуле ДНК во время репликации или рекомбинации, когда есть возможность смещения и неправильного спаривания нуклеотидов одной цепи с нуклеотидами другой. Считается, что встраивание молекул акридиновых красителей между неправильно спаренными основаниями стабилизирует такие измененные структуры. Если это так, то вероятность сохранения неправильно спаренной конфигурации нуклеотидов во время синтеза ДНК и воссоединения брешей довольно высока, что обусловливает инсерцию или делецию оснований на одной из цепей.