

Транспозоны:
3. Если Ds переносится внутрь гена W, то ген подавляется: нет производства антоцианов. Впоследствии, Ds может выскочить из гена, и экспрессия дикого типа гена W восстанавливается.
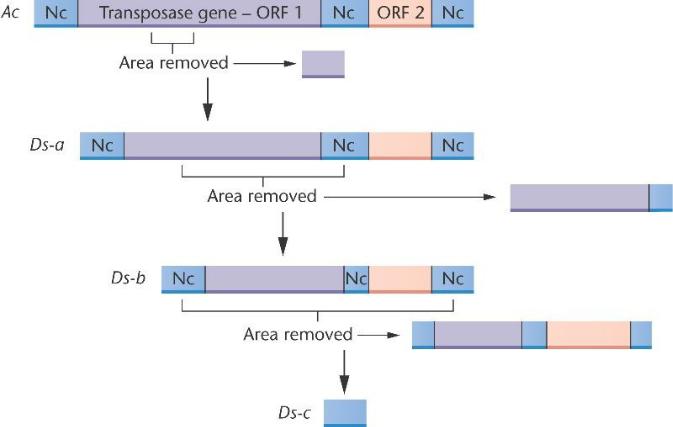
Сравнение структуры Ac и Ds элементов.
Последовательность Ac содержит некодирующие регионы (Nc) и открытые рамки считывания (ORF), которые кодируют фермент транспозазу и обеспечивает последовательность способностью "прыгать". Nc, также содержащие инвертированные терминальные повторы, здесь не показаны. Ds элементы похожи на Ac, но содержат делецию в гене транспозазы, что делает их зависимыми от Ac элементов для транспонирования.
Copia и Р-элементы у дрозофилы
В 1975 г. у дрозофилы идентифицировали группу генов, названных copia, с которых транскрибировалось множество копий РНК (отсюда название). Число этих генов в геноме до 30, они близки по структуре, их последовательности почти одинаковы, они способны к транспозициям и распределены по всему геному.
Гены этого семейства – только один пример из почти 30 семейств мобильных элементов дрозофилы, каждое из которых представлено 20-50 копиями генов. В сумме эти элементы составляют 5% генома дрозофилы и более половины средних повторов ДНК. Считается, что около половины видимых мутаций у дрозофилы обусловлено инсерциями транспозонов в гены дикого типа.
Несмотря на вариабельность последовательностей у членов разных семейств мобильных элементов, они имеют общую структуру, что связано с инсерцией и эксцизией в процессе транспозиции. Каждый ген copia содержит примерно 5000-8000 п.н., включая прямой концевой повтор 276 п.н. В пределах каждого повтора имеется короткий инвертированный концевой повтор длиной 17 п.н.
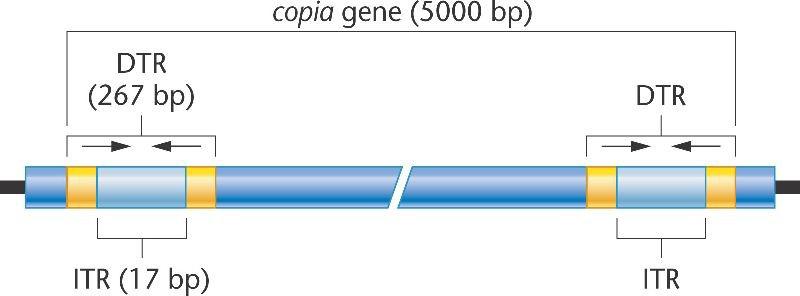
Каждый элемент copia (транскрибируется в "обильные" количества РНК) состоит из перевернутого концевого повтора (ITR) с последовательностью длинного прямого концевого повтора (DTR) на каждом конце.
Другая группа мобильных элементов дрозофилы – семейство Р- элементов. Это семейство открыли при исследованиях гибридного дисгенеза, состояния при котором мухи частично стерильны, у них высока частота мутаций и хромосомные аберрации в потомстве от скрещивания с некоторыми линиями дрозофилы.
Гибридный дисгенез обусловлен частыми транспозициями Р-элемента в клетках зародышевой линии. Встраиваясь в область гена или вблизи его, Р-элемент нарушает его экспрессию и вызывает мутации.
Последовательность Р-элемента содержит 2900 п.н., включая инвертированный концевой повтор длиной 31 п.н. Она кодирует не менее 2 белков, один из которых транспозаза, необходимый для транспозиций Р-элемента. Этот фермент продуцируется только в клетках зародышевой линии, поэтому транспозиции Р-элемента тканеспецифичны.
Мутации появляются в тех случаях, когда Р-элемент встраивается в кодирующую последовательность гена и нарушает его экспрессию. При его инсерции в промоторную область изменяется уровень экспрессии гена. Инсерции и интроны могут нарушить сплайсинг или прервать транскрипцию ДНК.
Мобильные элементы у человека
В геноме человека мобильные элементы представлены Alu- семейством коротких диспергированных повторов или SINE средней длины. Геном содержит около 500 тыс. копий этих последовательностей длиной 200-300 п.н.
Члены Alu-семейства считаются мобильными элементами по нескольким причинам. Наиболее важно, что их последовательности длиной 200-300 п.н. с каждой стороны граничат с повторами длиной 7-20 п.н., сходными с IS- элементами. Эти повторы связаны с инсерцией последовательностей в процессе транспозиции. Структура кластеров Alu-последовательностей в разных тканях одного организма, у здоровых и больных варьирует.
Были обнаружены и внехромосомные копии этих последовательностей.
Процессированные псевдогены
Процессированные псевдогены или ретропсевдогены - это ретропоследовательности, которые потеряли свою функцию. Они несут все признаки функциональных ретропоследовательностей, но имеют молекулярные дефекты, которые не дают им экспрессироваться.
Известны псевдогены, ведущие свое происхождение от всех типов РНК (мРНК, тРНК, рРНК, мя-РНК). Псевдогены транспортных РНК особенно интересны, поскольку они являются наиболее убедительным доказательством того, что процессированные псевдогены действительно образуются путем обратной транскрипции. Все ядерные тРНК имеют ССА последовательность на 3'-конце, которая не кодируется генами тРНК, а добавляется в результате посттранскрипционной ферментативной модификации. Процессированные же псевдогены тРНК часто содержат на 3'-конце ССА последовательность.
В среднем число процессированных псевдогенов больше, чем предковых функциональных генов. В некоторых случаях число процессированных псевдогенов может превосходить число соответствующих функциональных генов на несколько порядков.
Одним из примеров является семейство Alu-повторов.