

Опыты по трансформации
В 1927г. Ф. Гриффит проводил опыты со штаммами возбудителей стрептококковой пневмонии – вирулентными (вызывали заболевание) и невирулентными. Первые имели полисахаридную капсулу, вторые – нет
(разрушались иммунными клетками).
Убитые нагревом вирулентеые клетки не вызывают болезни, Смесь живых невирулентных и убитых вирулентных клеток неожиданно вызвала заболевание у мышей, в их крови обнаружили живые вирулентные клетки, идентичные убитым клеткам. Невирулентные клетки не мутировали (контрольные мыши не заболевали), но живые невирулентные и убитые вирулентные взаимодействовали между собой.
Превращение невирулентных клеток в вирулентные с помощью убитых клеток – трансформация. Ф. Гриффит считал, что трансформирующим фактором служит часть капсулы.

Вирулентные штаммы Diplococcus pneumoniae обладают капсулой, которая
делает их устойчивыми к иммунной системе животного-хозяина, вызывая фатальную пневмонию. Вирулентный штамм формирует гладкие (S) колонии в культуре, и его можно отличить от авирулентного,
формирующего шероховатые (R) колонии.

Гриффит показал, что когда мышам вводили смесь живых авирулентных бактерий, смешанных с убитых нагреванием вирулентными клетками, у мышей развилась пневмония, и они умерли, причем живые вирулентные
бактерии были обнаружены в крови.
Авирулентные бактерии были преобразованы в вирулентные.
Почему Гриффит не восстановил авирулентные бактерии из мертвых мышей?
Авирулентные бактерии были уничтожены фагоцитарными клетками
иммунной системы хозяина. Тем не менее, небольшой процент был трансформирован; они размножились и в конце концов убили хозяина.
Далее было показано, что этот процесс происходит и in vitro. Эвери, Маклеод и Маккарти доказали, что трансформирующим фактором является ДНК. Это было показано в опыте с использованием трех ферментов – протеазы, рибонуклеазы и дезоксирибонуклеазы, разрушающих соответственно белки, РНК и ДНК, причем только в последнем случае трансформирующая активность исчезала. Трансформирующий фактор взаимодействует с невирулентными клетками, включая ряд метаболических реакций, приводящих к синтезу полисахарида капсулы.
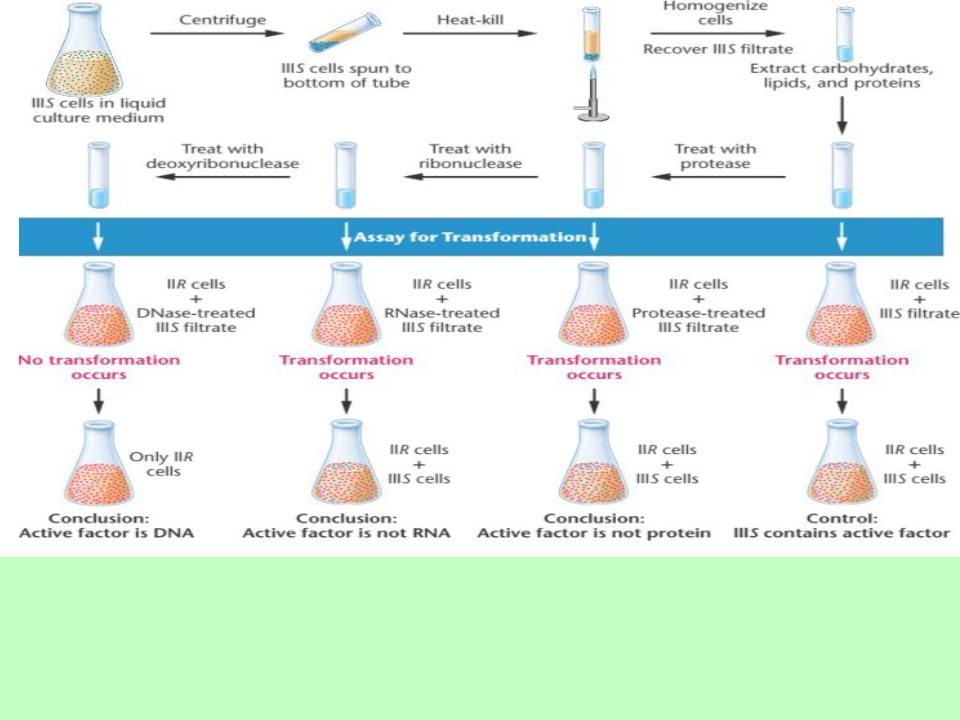
Эйвери, Маклеод и Маккарти изолировали трансформирующий фактор путем центрифугирования, чтобы собрать клетки, а затем убить их нагревом и извлечь макромолекулы.
Этот фильтрат способен к трансформации.
Инактивация или удаление полисахаридов, белков и РНК - фильтрат способен к трансформации. Добавление ДНКазы (фермента, расщепляющего ДНК) к фильтрату прекращает его трансформирующую активность, доказывая, что "активный фактор» - это ДНК.

Т-четный фаг, такой как T2 и T4, адсорбируется на клеточной стенке своего хозяина и внедряет свой генетический материал в клетку. Этот материал
направляет воспроизводство большого количества фагов, в конечном итоге лизируя хозяйскую клетку и выпуская свое потомство в литическом цикле.
Эксперимент Херши-Чейз
Другие доказательства роли ДНК как переносчика
генетической информации – при исследовании бактериофага Т2, инфицирующего E. coli.
Было известно, что фаги Т2 состоят наполовину из белков, наполовину из ДНК. В эксперименте А. Херши и М. Чейз метили белки радиоактивной серой, ДНК – радиоактивным фосфором, поскольку белки не содержат фосфора и ДНК серы. Оказалось, что при лизисе инфицированных меченым фагом бактерий освобождалось потомство, меченное фосфором, т.е. белковая оболочка фага остается снаружи бактериальной клетки-хозяина, а для образования вирусных частиц важна только вирусная ДНК
– генетический материал фага Т2.
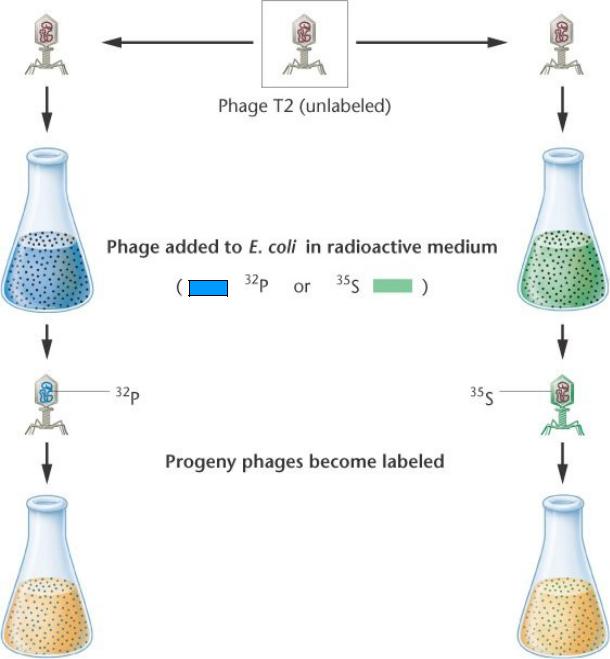
Hershey (Nobel 1969) и Chase
использовали радиоизотопы для мечения T2 фагов.
Нуклеотиды содержат
фосфор (P), поэтому 32P метит DNA в
головке фага.
Некоторые
аминокислоты
содержат серу (S),
поэтому 35S метит белковую оболочку.
Различно меченные фаги использовали для заражения немеченных бактерий.

После заражения, встряхиванием отделяют меченые фаговые оболочки от бактерий и разделяют оба компонента центрифугированием.
35S-меченный белок оставался вне клетки в составе фаговой оболочки («тени"). 32P-меченная ДНК была обнаружена в клетках, которые в конечном итоге лизируются, продуцируя потомство 32P-меченных фагов.
Прямые и непрямые доказательства значения ДНК у эукариот
В 50х гг эксперименты с ДНК – только на прокариотах, но предполагалось универсальность генетического материала для всех организмов
Непрямое доказательство – распределение ДНК
Предполагалась корреляция между плоидностью клеток (n, 2n ) и количеством молекул-носителей генетического материала. Действительно, в диплоидных клетках по сравнению с гаплоидными примерно вдвое больше ДНК, для белков такой корреляции нет