

Мутации
Новые аллели в популяции возникают только благодаря мутациям. Мутации происходят случайным образом, они не направлены на преимущества или недостатки организма.
Чтобы установить, являются ли мутации существенной движущей силой, изменяющей частоты аллелей, нужно измерить частоту возникновения мутаций.
Большинство мутаций рецессивно, поэтому для оценки частоты мутагенеза используют косвенные методы теории вероятностей и статистики или крупномасштабные исследования популяций.
Прямые методы анализа можно использовать только для достоверно доминантных мутаций.
Для точности результатов необходимо, чтобы аллель определял четко различимый фенотип, отличный от фенотипов с рецессивными аллелями, чтобы признак проявлялся полностью и чтобы была исключена возможность возникновения идентичного фенотипа за счет действия негенетических факторов.
Скорость мутагенеза – отношение числа новых мутантных аллелей к определенному числу гамет.
Зная скорость мутагенеза, можно оценить, в какой степени мутации могут привести к изменениям частот аллелей при смене поколений.
Пусть мутация возникает как доминантная у 2 из 100000 новорожденных. Родители фенотипически нормальны. Зигота имеет две копии каждого гена – значит на практике как бы изучение 200000 гамет.
Если предположить, что особи-мутанты гетерозиготны, то обнаружено два мутантных аллеля на 200000 копий гена, т.е.
скорость мутации = 2/200000 = 1х10-5.
Зная скорость мутагенеза, можно оценить в какой степени мутации могут привести к изменениям частот аллелей при смене поколений
В популяции из 500000 человек у всех генотип dd. Первоначальная частота аллеля d = 1, аллеля D = 0. Генофонд состоит из 1 млн гамет с аллелем d. При этом 1.4 на каждые 100000 аллелей мутируют с образованием аллеля D.
Теперь частота аллеля D = (1000000-14)/1000000 = 0.000014, т.е. мутациям самим по себе нужно очень много времени для заметного сдвига частот в популяции
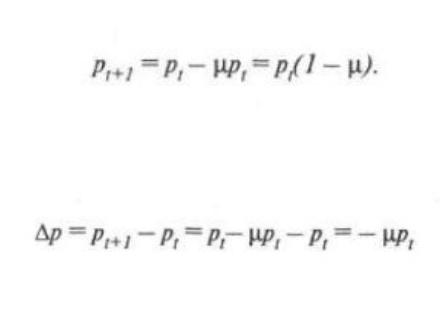
Мутационный процесс в популяции.
Частота спонтанных мутаций низка, поэтому изменение частоты аллеля только в результате мутационного процесса происходит чрезвычайно медленно.
Обозначим частоту аллеля А в поколении t как рt и частоту мутаций А -> а за
поколение — u.
Тогда частота аллеля А в следующем t + 1 поколении будет равна
Изменение частоты аллеля за одно поколение составит
В общем случае, имея в поколении g два аллеля А с частотой р и а с частотой q и скоростью мутагенеза µ, приводящего к превращению аллеля А в аллель а, можно рассчитать частоты аллелей в следующем поколении:
pq+1 = pg - µpg qg+1 = qg + µpg
Скорость замены аллеля А на аллель а при первоначальной частоте аллеля А = 1 и скорости мутагенеза 1х10-5 упадет до 0.5 за 70000 поколений.
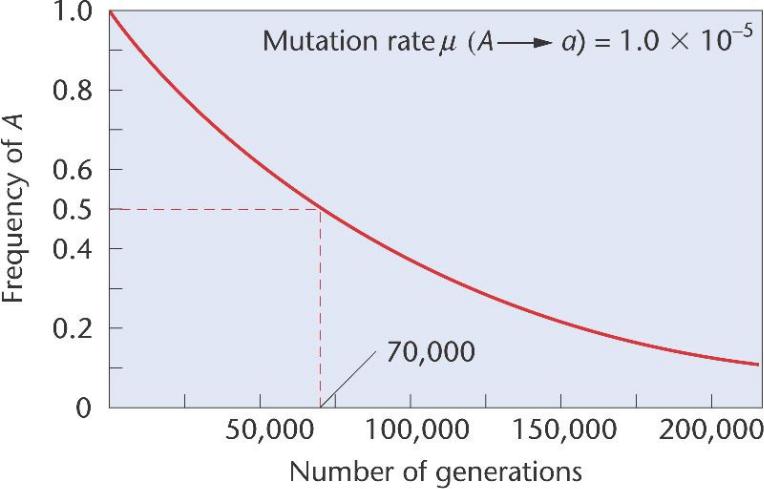
При скорости мутации (μ) 1.0 x 10-5, требуется 70,000
поколений для уменьшения частоты аллеля A от 1.0 до 0.5.
Таким образом, мутация сама по себе играет относительно несущественную роль в изменении частоты аллеля.
Мутагенез создает материал для эволюции, но частота аллелей, возникших при мутации, обычно определяется естественным отбором или дрейфом генов.
Частота мутантного аллеля, вызывающего муковисцидоз (рецессивно-аутосомный признак) в Европе 2%. Существует сильный отбор против рецессивных гомозигот.
Какие факторы удерживают частоту аллеля на уровне 2%?
Основная гипотеза – гипотеза превосходства гетерозигот, по которой отбор против рецессивных гомозигот уравновешивается отбором в пользу гетерозигот. Есть предположения, что гетерозиготность по этому гену повышает устойчивость к брюшному тифу
