

Генетические исследования бактериофагов
Бактериофаги или фаги – это бактериальные вирусы, которые могут участвовть в передаче генетической информации путем трансдукции.
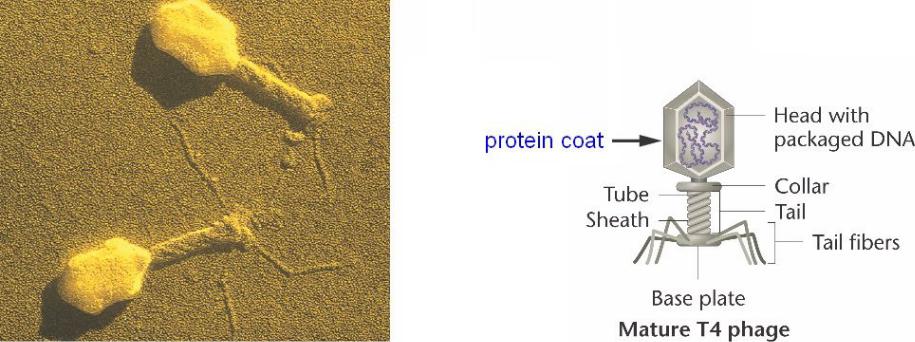
Бактериофаг T4 – один из группы литических (или вирулентных)
бактериальных вирусов, называемых T-четными фагами. «Головка" состоит из белковой оболочки, содержащей ДНК. «Хвост" содержит
воротник и сократительную оболочку, окружающую центральное ядро;
хвостовые фибриллы, выступающие за базальную пластинку,
содержат сайты связывания, которые узнают клеточную стенку E. coli.
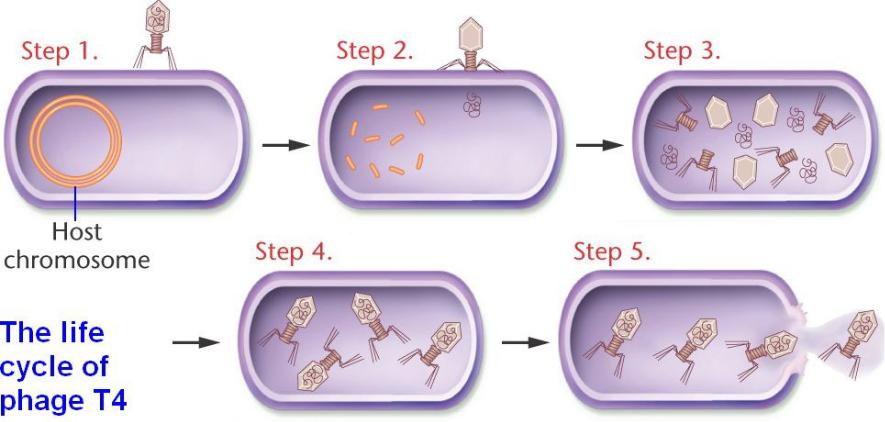
1)Вирус связывается с поверхностью бактериальной клетки-хозяина хвостовыми фибриллами.
2)Оболочка хвоста сжимается и происходит проникновение центральной части
сквозь клеточную стенку.
3)Белковая оболочка остается вне хозяина, в то время как вирусная ДНК впрыскивается и ДНК хозяина деградирует.
4)Молекулы вируса синтезируются с использованием ресурсов хозяина; и
начинается сборка потомства фага из компонентов (~ 200 частиц).
5) Фермент фага (лизоцим) разрывает (лизирует) клетку, освобождая фаговые частицы и завершая литический цикл.
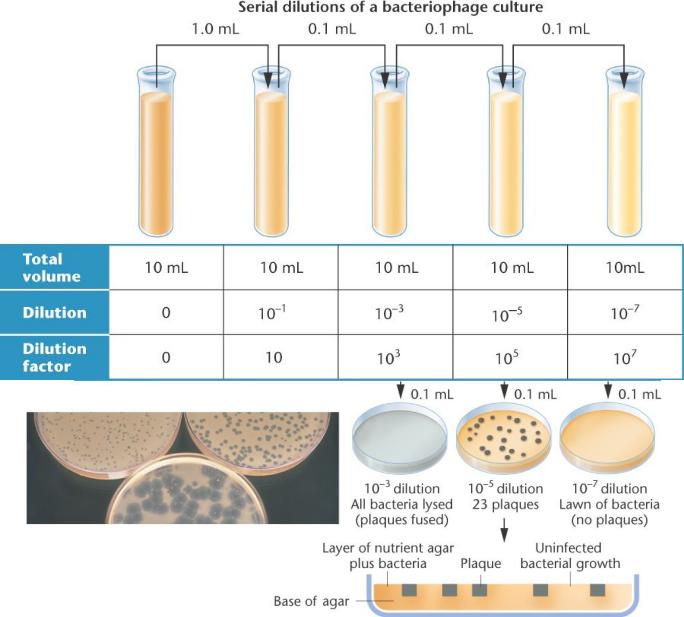
Метод бляшек
концентрированной
культуры
бактериофагов начинается с серийных разведений вирусной культуры. Небольшой образец затем смешивали с бактерией-хозяином и высевали на питательный агар. После инкубации бактерии растут в форме "газона" клеток,
с четкими областями ("бляшками"),
указывающими, где один фаг изначально заразил одну бактерию.
Здесь 23 фаговые бляшки, полученных из 0.1 ml аликвоты (образца) разведением |
10– |
5. Для расчета начальной неразведенной вирусной плотности (частицы / ml): (23 * 105 ) / 0.1 ml = 23 * 106 / ml, или 2.3 * 107 / ml
Лизогения
Взаимодействие бактериофагов и бактерий не всегда приводит к размножению фага и лизису бактериальных клеток. Вирус может существовать в своеобразном симбиозе с бактерией.
Молекулярные основы такого симбиоза сейчас понятны. После проникновения в клетку бактерии вирусная ДНК интегрируется в бактериальную хромосому и наступает стадия лизогении. В результате вирусная ДНК реплицируется в составе бактериальной хромосомы и попадает в каждую из дочерних клеток, образующихся в результате деления инфицированной бактерии. При этом не происходит лизиса клеток и выхода из них зрелых вирусных частиц. Однако при определенных воздействиях (УФ, химическое вещество ) вирусная ДНК вырезается из хромосомы, начинает автономно реплицироваться и после сборки вирусных частиц происходит лизис бактериальной клетки
Вирусная ДНК интегрирует в хромосому бактерии в виде профага Вирусы, которые могут лизировать бактерии, а также
встраиваться в их хромосомы, называются умеренными, а вирусы, которые всегда лизируют бактериальные клетки – вирулентными Бактерии, содержащие профаги, называются лизогенными, они могут лизировать в результате индукции профага.
Вирусная ДНК, реплицирующаяся либо в составе бактериальной хромосомы, либо автономно, в цитоплазме клетки, относится к эписомам.
Трансдукция – перенос бактериальной ДНК вирусом
Зиндер и Ледерберг в 1952 г. после смешения двух разных ауксотрофных штаммов бактерий сальмонеллы обнаружили прототрофные клетки, но оказалось, что рекомбинация между бактериями по своему механизму отличается от переноса
F фактора у E coli. Открытая рекомбинация у бактерий была обусловлена бактериофагами и получила название -трансдукция

Трансдукция.
Zinder и Lederberg (Nobel 1958) поместили два
ауксотрофных штамма
Salmonella в противоположные части U-образной трубки
Дэвиса.
Прототрофы обнаруживались со стороны, содержащей клетки
LA-22, но не со стороны с клетками LA-2.
Добавление ДНКазы, фермента, расщепляющего
ДНК, не препятствует рекомбинации, поэтому это не
трансформация.
Фильтр препятствует контакту
клеток, поэтому это не
конъюгация.
Рекомбинация прекращается, когда размер пор в трубке меньше размера фагов.
o
Природа трансдукции
Позже трансдуцирующие фаги были обнаружены у других бактерий. Существуют различные виды трансдукции. Помимо трансдукции умеренными фагами лизогенных бактерий возможна трансдукция во время обычного литического цикла.
Иногда небольшой участок бактериальной ДНК пакуется вместе с вирусной ДНК и, поэтому трансдуцирующий фаг содержит как вирусные гены, так и несколько бактериальных генов. Однако в случаях, когда упаковывается только бактериальная ДНК, в вирусной головке содержится около 1% длины бактериальной хромосомы. В любом случае способность к инфекции не связана с типом ДНК, упакованной в головке трансдуцирующего фага
Когда фагом инъецируется не вирусная, а бактериальная ДНК, она либо остается в цитоплазме, либо рекомбинирует с гомологичным участком хромосомы инфицированной клетки. В первом случае ДНК случайным образом попадает в одну из дочерних клеток, т.е. частично диплоидной становится только одна клетка – абортивная трансдукция
При рекомбинации с гомологичным участком бактериальной хромосомы во втором случае, трансдуцированные гены реплицируются в составе этой хромосомы и передаются во все дочерние клетки – полная трансдукция
Абортивная и полная трансдукции – частные случаи генерализованной трансдукции, для которой характерен случайный перенос фрагментов ДНК и генов. Существует вероятность упаковки в головку фага любого из фрагментов бактериальной ДНК.
В большинстве случаев генерализованная трансдукция проходит по типу абортивной, полная на порядок реже