

Подготовка у универсиаде 2012 / Генетика (Жимулев) / 8-2ver7
.pdfМолекулярныеГлава8 |
механизмы мутагенеза... |
Молекулярные механизмы |
... |
|
мутагенезаГлав 8 |
||
|
|
|
|
событие, действуют только на особую пару последовательностей-мишеней.
3.Рекомбинация между негомологичными последовательностями нуклеотидов происходит в клетках прокариот и дрожжей достаточно редко, а в клетках млекопитающих – довольно часто. К негомологичной рекомбинации можно отнести процесс случайного встраивания вирусной или плазмидной ДНК в ДНК
клеток животных.
Ниже проведено более подробное описание этих процессов.
8.3.1. Гомологичная рекомбинация
Гомологичнаярекомбинацияпроисходит междудвумядуплекснымимолекуламиДНК. Следует подчеркнуть, что ферменты, участвующие в этом процессе, могут использоватьвкачествесубстраталюбуюпару гомологичныхпоследовательностей.
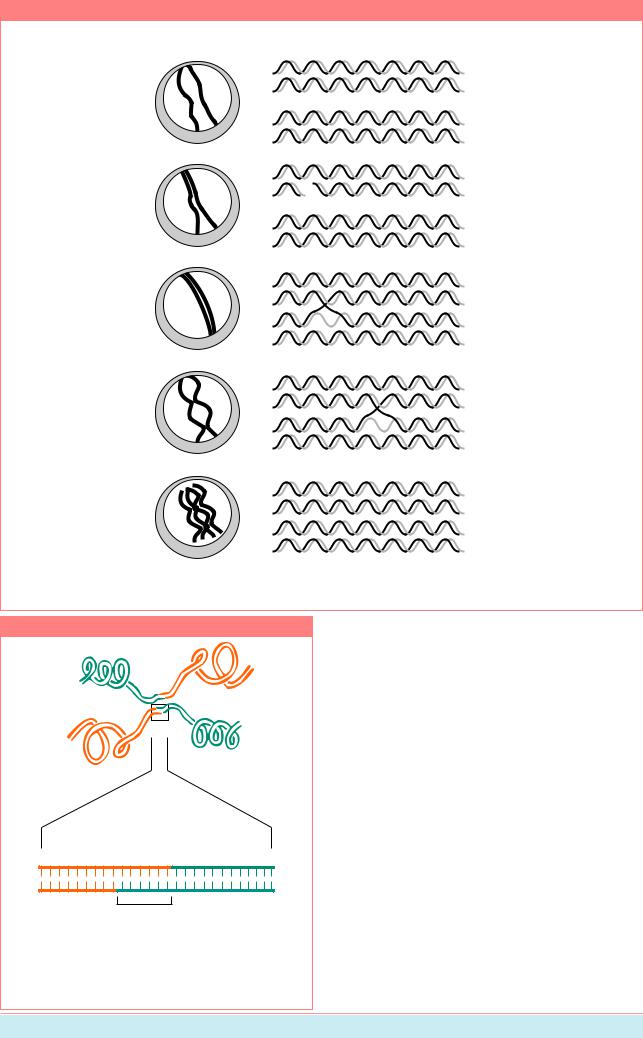
НаРис.8.17показанасхемапрохождения стадий мейоза и сопутствующих им этапов осуществления кроссинговера на молекулярном уровне. Началом профазы мейоза можно считать стадию, на которой становятся видимыми индивидуальные хромосомы.ВкаждойизэтиххромосомДНК ужереплицированаисодержитдвесестринские хроматиды, состоящие из дуплексной ДНК. Гомологичные хромосомы притягиваются другкдругу, коньюгируютводномилиболее районов,формируябиваленты.Когдапроцесс спаривания хромосом завершен, хромосомы соединяются латерально за счет структуры, называемойсинаптонемальнымкомплексом.
Рекомбинация между хромосомами подразумевает физический обмен частями, происходящий по принципу “разрыв и воссоединение”, в ходе которых две несестринские хроматиды рвутся и затем воссоединяются.Когдахромосомыначинают расходиться, их контакты между собой остаются видимыми в виде так называемых хиазм.
Традиционнойточкойзренияявляетсято, что хиазмы представляют собой отражение
существования кроссинговера, хотя формальных доказательств этой связи до сих порнеполучено.
По-видимому, первым шагом к началу рекомбинацииДНКявляетсясближениедвух дуплексныхмолекулДНК.
Для обмена между дуплексами ДНК необходимообразованиепокрайнеймередвух брешей-ников. В механизме общей рекомбинации образование этих ников и запечатывание разрывов скоординированы таким образом, что они встречаются только тогда, когда две спирали ДНК имеют обширные районы гомологии. Для того, чтобыполучитьструктуру, изображеннуюна Рис. 8.18, необходимо, чтобы в каждой из четырех цепей образовалась брешь с последующим лигированием.
Существует достаточно доказательств, что даже единственная брешь только в одной цепи молекулы ДНК достаточно для инициацииобщейрекомбинации.Химические препараты или излучение, приводящие к образованиям однонитчатых ников, будут стимулировать события рекомбинации. Кроме этого, существует особый белок – RecBCD ó E.coli – который делает однонитчатые ники в молекулах ДНК. Этот белок также обладает геликазной активностью, он гидролизует АТФ и перемещается вдоль по молекуле ДНК. За счет комбинации нуклеазной и геликазной активностей белок RecBCD создает в молекуле ДНК однонитчатый “усик” (Рис. 8.19). На Рис. 8.20 показано как “усик” может инициировать взаимодействие между двумя комплементарными нитями ДНК. Процесс, однако, более сложен. Для того, чтобы цепь из одной гетеродуплексной молекулы смогла внедриться в другую двухцепочечную ДНК, у E.coli функционирует белок RecA, продуцируемый геном recA. Белок RecA в виде больших кооперативных кластеров плотно связывается с однонитчатой ДНК, образуя нуклеопротеиновый филамент. Этот филамент обладает несколькими особенностями. Поскольку в нем несколько сайтов связывания с ДНК, то есть
|
|
218 |
218 |
|
|
...Молекулярные механизмы мутагенеза |
Глава 8 |
|
|
Рисунок 8.17
Стадии мейоза
Лептотена
Конденсированные
хромосомы становятся видимыми, часто прикреплены к ядерной мембране
Зиготена
Хромосомы начинают спариваться в отдельных районах
Пахитена
Синаптонемный комплекс образуетя по всей длине спаренных хромосомы
Диплотена
Хромосомы
разделяются, но соединены хиазмами
Диакинез
Хромосомы
конденсируются. отделяются от мембраны, но хиазмы остаются. Все 4 хроматиды становятся видимыми
Молекулярные события
Каждая хромосома реплицирована и состоит из двух сестринских хроматид
Инициация
В одной хромосоме возникает разрыв
Обмен цепей
Одноцепочечный обмен с другой хромосомой, образование гетеродуплекса
Ассимиляция
Участок формирования гетеродуплекса увеличивается
Разделение
Хромосомы разделяются с помощью брешей - ников
События мейоза и обмена цепей в молекулах ДНК (Из: Lewin, 1994, P. 968).
Рисунок 8.18 |
Участок сочленения |
Сочленение (joint) – объединяет две молекулы ДНК |
в том месте, в котором произошел кроссинговер. |
Участок сочленения может иметь длину в |
несколько тысяч нуклеотидов. (Из: Alberts et al., |
1994, P. 264). |
возможность держать одиночную и двойную цепи ДНК вместе. Эти сайты позволяют белку RecA катализировать многоступенчатую реакцию (называемую синапсисом) между двуцепочечной молекулой ДНК и гомологичными районами одноцепочечной.
Первым шагом в синапсисе является гомологичное спаривание оснований между комплементарными последовательностями нуклеотидов. В результате образуется трехцепочечная структура. После этого короткий гетеродуплексный участок, в котором нити из двух различных молекул начинали спариваться, увеличивается из-за “миграции ветви” (Ðèñ.8.21). “Миграция ветви” может происходить в любой точке, где две одиночных цепи ДНК, имеющих те же самые последовательности, конкурируют
219

Глава 8
Рисунок 8.19
RecBCD
белок
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I
|
|
|
|
|
|
|
|
Присоединение к концу |
|||||||||||
Сайт узнавания |
|
|
|
|
двойной цепи и |
|
|
||||||||||||
|
I |
I I |
I |
|
|
|
|
передвижение по ней |
|||||||||||
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I I I I I |
|
|
I I I I I I I I I I I I I I I I I I I I I |
||||||||||||||||
I I I I I |
I |
|
II I I I I I I I I I I I I I I I I I I I I |
||||||||||||||||
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
I I |
|
|
|
|
|
Одноцепочечный разрыв в |
|||||||||||
Перемещающаяся |
|
|
|
|
|||||||||||||||
|
|
|
|
сайте узнавания |
|
|
|||||||||||||
петля одноцепочечной |
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
ÄÍÊ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I |
|||||||||||||||||||
I I I I I I I I I I I I I I |
|
|
I |
I |
I |
|
I I I I I I I I I I I I I |
||||||||||||
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
|
|
Перемещение |
|
|
|
|||||||||
|
|
|
|
|
|
|
|
однонитевого “усика” |
|||||||||||
|
|
||||||||||||||||||
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I |
|||||||||||||||||||
I I I I I I I I I I I I I I |
|
|
|
|
|
|
|
|
|
|
I |
I |
I |
I |
I |
I I I I I |
|||
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
I |
I |
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
3’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Îäèí |
|
èç |
вариантов |
|
|
|
инициации |
||||||||||||
кроссинговера. У E.coli белок RecBCD необходим для осуществления общей рекомбинации. Белок связывается с концом спирали ДНК и затем, используя энергию от гидролиза связанной с ним молекулы АТФ, продвигается вдоль молекулы со скоростью 300 нуклеотидов в секунду. Особый сайт опознавания (последовательность ДНК из 8 нуклеотидов, встречающаяся во многих районах хромосомы E.coli) разрезается в пределах “мигрирующей петли”, создаваемой белком RecBCD, в результате чего образуется однонитчатый “усик”, который, как полагают, инициирует генетическую рекомбинацию, спариваясь с гомологичной ДНК (Из: Alberts et al., 1994, P. 264).
за возможность спариваться с одной и той же комплементарной цепью. Неспаренный участок одной из одиночных цепей заменяется спаренным районом другой одиночной цепи, двигая точку ветвления. Спонтанное движение ветви равновероятно влюбомнаправлении(Рис.8.21а).Поскольку RecA белок катализирует однонаправленное движение ветви, это создает район гетеродуплексадлинойвнесколькотысячпар оснований (Рис. 8.21б).
После этой стадии у большинства
Молекулярные механизмы мутагенеза...
изученных организмов наступает стадия формирования перекрестного “обмена цепей” или “структур Холлидея” (Рис. 8.22).
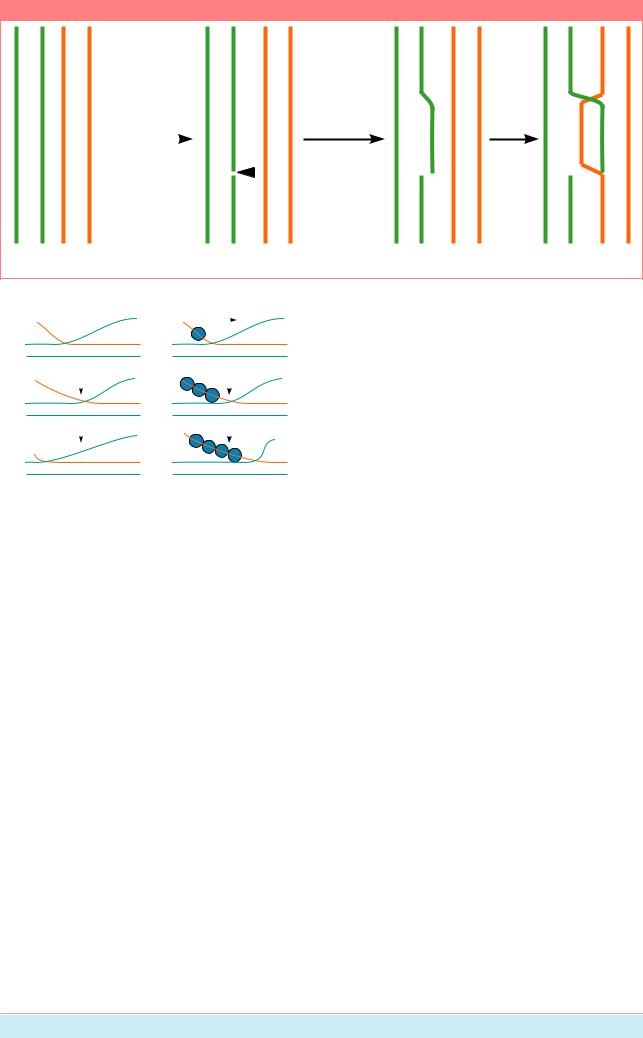
В этих структурах две гомологичные молекулы ДНК, которые раньше были спарены,теперьудерживаютсявместезасчет сформировавшихся обменов между двумя из четырехцепей:пооднойизкаждоймолекулы ДНК. Структура Холлидея имеет две особенности: 1. Точка обмена между цепями (см. Рис. 8.22) может быстро мигрировать вперед и назад. 2. Она состоит из двух пар цепей:однапарапересекающихсяиоднапара непересекающихся. Может происходить изомеризация: после серии вращений молекулы исходно непересекающиеся, становятся пересекающимися, и наоборот. Изомеризацияможетпроисходитьспонтанно ипод контролем клетки (Рис. 8.23).
Для того, чтобы восстановить две отдельные спирали ДНК и т.о. закончить процесс спаривания молекул, две пересекающиесяцепидолжныбытьразрезаны. Если они разрезаны до изомеризации, две исходныеспиралиотделяютсяоднаотдругой почти неизмененными, только с обменным очень коротким фрагментом однонитчатой ДНК. Если пересекающиеся нити разрезаны после изомеризации, одна секция каждой из исходныхспиралейДНКсоединяетсяссекцией другой молекулы, другими словами, две спиралиДНКиспытываюткроссинговер.
Если изомеризация не произошла, разрезание двух пересекающихся цепей просто бы прекратило обмен нитей и кроссинговер не произошел бы.
Если изомеризация происходит, разрезание двух пересекающихся цепей создает две молекулы ДНК, которые испытывает кроссинговер. Таким образом, изомеризация, как полагают, необходима для разрываивоссоединениядвухгомологичных спиралей ДНК, возникающих в результате общей генетической рекомбинации (этап A показан на Рис. 8.17).
220
...Молекулярные механизмы мутагенеза |
Глава 8 |
|
|
Рисунок 8.20
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I
IIIIIIIIIIIIIIIIОбразование |
I I I I I I I I I I I I I I I I |
|
одноцепочечного |
||
I I |
I I |
|
IIIIразрыва |
I I |
|
+II |
|
I I I I |
|
||
I I I I I I I I I I I I I I I I I I I I |
I I I I I I I I I I I I I I I I I I I I |
|
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I
Образование
одиночной нити ДНК
IIII IIII IIII IIII IIII IIII IIIII II IIIII II IIIIII II IIIIII II IIIIII IIII IIII IIII IIII IIII IIII II II
Обмен
цепями
ÄÍÊ
I I I I I I I I I I I I I I I |
IIIIIIIIIIIIIIII |
IIIIII IIIIII IIIIIIIIIIII |
|
I |
II |
I I I I I I I I I I I I I I I I I I I I I I I I I I I I |
|
Первоначальный обмен нитями в схеме общей рекомбинации (Из: Alberts et al., 1994, 265).
Рисунок 8.21 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Сайт-специфическая рекомбинация |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
5’ |
|
|
|
Движение белков |
|
|
|
I 3’ |
|
была открыта в результате исследований |
|||||||||||||||||||||||||||||
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
II I I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
II I |
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
механизма перемещений бактериофага λ по |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
I |
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
5’ |
|
|
|
|
|
I |
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
3’ |
5’ |
|
|
|
|
|
I |
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
|||||||||||
|
|
|
|
|
IIIIIII |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IIIIIII |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||
|
|
I I I I I I |
I |
I |
|
|
I I I I I I I I I I I I I I I |
|
|
I I I I I I |
I |
I |
|
|
|
I I I I I I I I I I I I I I I |
|
|
хромосоме E.coli. В интегрированном |
||||||||||||||||||||||||||||||||||||||||||||||
3’ |
I I I I I I I I I I I I I I I I I I I I I I I I I |
5’ |
3’ |
I I I I I I I I I I I I I I I I I I I I I I I I I |
5’ |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
III I |
3’ |
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
III I |
3’ |
|
состоянии вирус внедрен в бактериальную |
||||||||||||
|
|
II |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
II |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
хромосому и реплицируется как часть ДНК |
|||||||||||
|
|
|
|
|
|
|
I |
I |
I |
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
I |
I |
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
3’ |
5’ |
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
|||||||||||||
|
I I I I I I I I I I |
I IIII |
|
I |
I I I I I I I I I |
I I I I I I I I I I |
IIII |
|
I |
I I I I I I I I I |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
I I |
|
|
I |
|
|
I I |
|
|
|
I |
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||
3’ |
I I I I I I I I I I I I I I I I I I I I I I I I I |
5’ |
3’ |
I I I I I I I I I I I I I I I I I I I I I I I I I |
5’ |
|
клетки-хозяина. Когда вирус проникает в |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
II I |
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
|
|
|
клетку, на |
матрице вирусного гена |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IIII |
|
|
|
|
|
|
|
|
|
II |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
||||||||||||
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
II |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
II |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
синтезируется фермент λ -интеграза. Этот |
|||||||||||
|
|
I |
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
5’ |
|
I |
|
|
|
II |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
I |
|
|
|
|
|
|
3’ |
|
|||||||||||||||
|
|
|
II |
I I |
I I I I I |
I |
I I I I |
I I I I I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
II |
I I I I I |
|
|||||||||||||||||||||||||||||||||||
|
|
I I I |
I I I |
|
I |
|
|
I I I I I I I I I I I I I I I I |
II |
|
I |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
3’ |
I I I I I I I I I I I I I I I I I I I I I I I I I |
5’ |
3’ |
I I I I I I I I I I I I I I I I I I I I I I I I I |
5’ |
|
фермент |
è |
катализирует |
процесс |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
а. Спонтанная миграция ветви |
|
|
б. Однонаправленная |
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
вперед и назад |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
миграция ветви, |
|
|
|
|
|
|
|
|
рекомбинации, начинающийся тогда, когда |
||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
контролируемая белками |
|
|||||||||||||||||||||||||||||||||
Два типа миграции ветви, наблюдаемые в |
|
несколько молекул белка интегразы плотно |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
экспериментах in vitro (Из: Alberts et al., 1994, |
|
связываются |
ñî |
специфическими |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
p. 267). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
последовательностями |
на кольцевой |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
хромосоме фага. Получившийся ДНК- |
|||||
8.3.2. Сайт-специфическая |
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
белковый комплекс теперь связывается со |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
рекомбинация |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
сходными, |
|
íî |
íå |
идентичными |
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
8.3.2.1. Характеристика |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
последовательностями на бактериальной |
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
хромосоме, |
приближая тем |
самым |
||||||||||||||||||||||||||||||||||||||||||||||
процесса |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
бактериальную и фаговую хромосомы (Рис. |
||||||||||||||||||
|
Вотличиеотобщейрекомбинации,сайт- |
|
8.24). Затем интеграза делает надрезы в |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
специфическаярекомбинацияпроисходитпод |
молекулах ДНК, формируя маленький |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
контролем |
|
|
|
|
|
ферментов, |
|
|
|
|
опознающих |
участок сочленения гетеродуплекса. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||
специфические |
|
последовательности |
ИнтегразанапоминаетДНК-топоизомеразув |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
нуклеотидов, присутствующие на одной или |
том отношении, что она формирует |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
двух рекомбинирующих молекулах. В этом |
ковалентную связь с ДНК там, где она ее |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
случае спаривание оснований между |
|
ðâåò. |
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
рекомбинирующими |
молекулами |
|
|
|
íå |
Тот же самый механизм сайт- |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
требуется, и даже когда оно происходит, |
специфической рекомбинации приходит в |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
сочленение гетеродуплексов имеет длину |
|
действие только в обратном направлении, |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
всеговнесколькопароснований.Спомощью |
когда фаг λ |
вырезается из его сайта |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
этого типа рекомбинации, бактериальные |
интеграции. |
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
вирусыимобильныеэлементыперемещаются |
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
по геному. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
221

Глава 8 |
...Молекулярные механизмы мутагенеза |
|
|
Рисунок 8.22
Две гомологичные спирали ДНК
Образование
одноцепочечных разрывов и обмен цепями
Образование
одноцепочечных разрывов и обмен цепями
Лигирование нитей с разрывами
Перекрестный обмен цепей ДНК (структуры Холлидея) (Из: Alberts et al., 1994, P. 268).
Рисунок 8.23
äâå
гомологичные
хромосомы
образование à крестообразной
структуры
á
Вращение в другой плоскости
â
 разрезание ã обеих
разрезание ã обеих
цепей ДНК
ä
äâå
хромосомы
после
кроссинговера
Изомеризация перекрестного обмена цепей (Из: Alberts et al., 1994, P. 269).
222

Молекулярные механизмы мутагенеза... Глава 8
Рисунок 8.24 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
повтор, - получаются делеция и дупликация |
||
|
|
кольцевая хромосома |
(Ðèñ. 8.25). |
||||||||||||||
бактериофага |
|
|
|
|
|
|
|
|
|
Всю систему можно ввести в геном |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
дрозофилы с помощью трансформации. Для |
|
|
|
|
|
|
|
|
специфические сайты |
этого создают две конструкции: в одной FLP |
||||||||
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
||||||||||
|
бактериальная |
|
|
присоединения |
ставится под промотор теплового шока, |
||||||||||||
хромосома |
|
|
|
|
|
бактериофага |
|
||||||||||
|
|
|
|
|
|
благодаря чему рекомбинацию можно |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
индуцировать, в другой конструкции между |
|
|
|
|
|
|
|
|
-интеграза |
|
|
FRT располагают ген white, по делеции или |
||||||
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
дупликации которого можно судить о |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
прошедшей рекомбинации (Рис. 8.26). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Очевидно, что если поместить FRT |
интеграза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
разрезает |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
последовательности в P-элемент, заставить |
||
ДНК хромосом |
|
|
|
|
|
|
|
|
|
|
|
|
|
его перемещаться по геному, можно |
|||
и соединяет |
|
|
|
|
|
|
|
|
|
||||||||
их заново |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
получить огромное количество линий с |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
инсерциями P-элемента, а следовательно и |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
FRT участков, в самые разнообразные |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
отсоединение |
|
участки генома дрозофилы. Затем отбирая |
|||||||
|
|
|
|
|
|
|
|
|
пары линий с известной локализацией P- |
||||||||
|
|
|
|
|
|
|
|
интегразы |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
элементов, можно направленно получать |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
делеции и дупликации между выбранными |
||||||||||||||
ДНК бактериофага, встроенная в |
|||||||||||||||||
бактериальную хромосому |
районами хромосом (Рис. 8.27). |
||||||||||||||||
Инсерция |
ÄÍÊ |
бактериофага |
λ |
â |
|
|
|
|
|
|
||||
бактериальную хромосому (Из: Alberts et al., |
8.3.3. Случайная рекомбинация |
|||||||||||||
1994, P. 271). |
|
|
|
|
|
|
|
|
Многиемобильныепоследовательности |
|||||
|
|
|
|
|
|
|
|
|
|
|||||
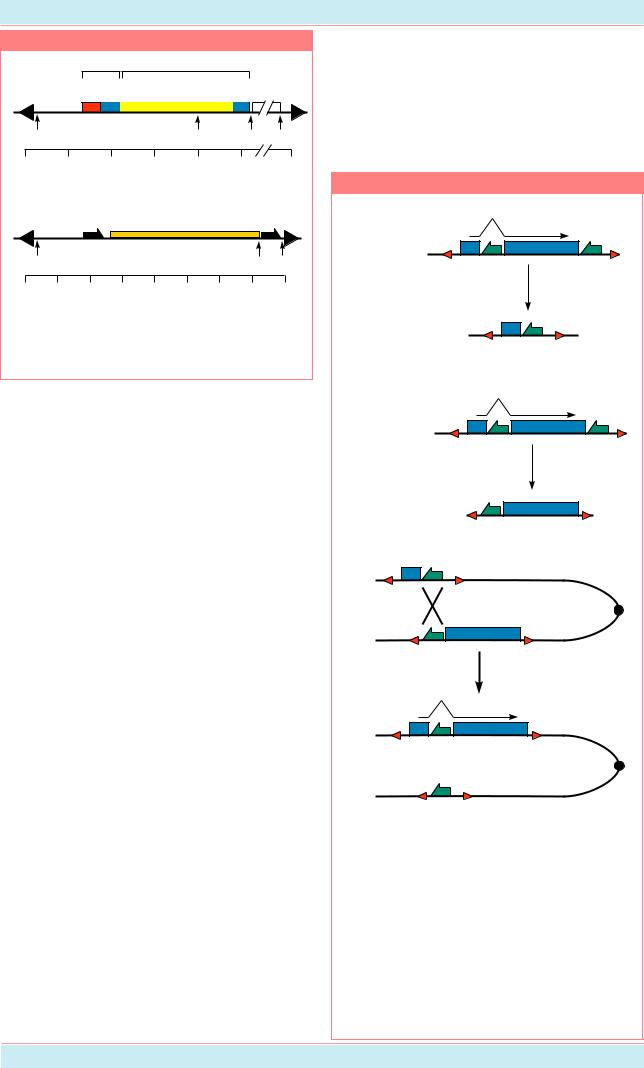
8.3.2.2. Экспериментальная |
|
ДНК,включаявирусыимобильныеэлементы, |
||||||||||||
сайт-специфическая |
|
|
|
кодируют интегразы (или по-другому, |
||||||||||
рекомбинация у дрозофилы |
|
транспозазы), которые помогают их ДНК |
||||||||||||
Система FLP из генома дрожжей |
встраиваться в хромосомы, используя |
|||||||||||||
содержит все необходимое (рекомбиназу) |
механизм, отличный от того, который |
|||||||||||||
для индукции рекомбинации. Сигнал от FLP |
используетсябактериофагомлямбда.Так же, |
|||||||||||||
воспринимается сайтами FRT (FLP Рисунок 8.25 |
|
|
|
|
||||||||||
recombination targets), имеющими à |
|
|
|
|
|
|
|
|||||||
длину |
около |
600 |
ï.í. |
FLP |
|
|
A |
FRT |
B |
A |
FRT |
C |
||
располагается в 2 m минихромосоме |
|
|
|
|
||||||||||
|
|
D |
|
C |
|
|
|
|||||||
(или плазмиде). Фрагменты FRT |
|
|
FRT |
D |
FRT |
B |
||||||||
|
|
|
|
|
|
|||||||||
могут находиться в любом участке |
|
|
|
|
|
|
|
|
||||||
генома. В плазмиде 2 m находится |
á |
|
|
|
|
|
|
|
||||||
какрекомбиназа(FLP), такипара FRT |
|
FRT |
|
|
FRT |
|
|
FRT |
||||||
|
|
|
|
|
|
|
|
|||||||
районов (Рис. 8.25). Этого набора |
|
|
|
|
|
|
|
FRT |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
генных элементов, заключенных в |
|
|
|
|
|
|
|
|
||||||
2мкм кольце ДНК, оказывается |
|
|
|
|
|
|
|
|
||||||
достаточно |
äëÿ |
прохождения |
|
|
|
|
|
|
|
|
||||
рекомбинации, при этом, если FRT- Результаты сайт-специфической рекомбинации в 2µ m |
||||||||||||||
последовательности ориентированы плазмиде |
дрожжей. |
Рекомбинация |
между |
|||||||||||
навстречу друг другу, то в результате инвертированными (а) и прямыми (б) повторами FRT. |
||||||||||||||
кроссинговера образуется инверсия, FRT – обозначены жирными полустрелками (Из: Golic, |
||||||||||||||
åñëè FRT расположены как прямой Lindquist, 1989, P. 500). |
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
223 |
Глава 8 |
|
|
|
|
|
|
|
|
Молекулярные механизмы мутагенеза... |
|||||||
Рисунок 8.26 |
|
|
|
|
|
|
|
Литература к разделу 8.3. |
||||||||
à |
|
hsp70 |
|
FLP |
|
|
|
|
Инге-Вечтомов С.Г. Генетика с основами |
|||||||
|
|
|
|
|
|
|
||||||||||
P |
промотор |
лидер |
кодирующая часть |
3’ |
ry+ |
P |
|
селекции. Москва, Высшая школа, 144- |
||||||||
|
|
|
|
168, 1989. |
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
Кушев |
Â.Â. |
Механизмы |
генетической |
|||
HindIII |
|
|
|
HindIII |
|
HindIII HindIII |
|
рекомбинации. Ленинград, Наука, 1-247, |
||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||
0 |
0.5 |
|
1.0 |
1.5 |
2.0 |
|
2.5 |
|
10 |
|
1971. |
|
|
|
||
á |
|
|
|
|
|
|
|
|
|
Рисунок 8.27 |
|
|
|
|||
|
|
|
|
|
|
|
|
|
à |
|
|
|
|
|
||
P |
|
FRT |
|
w hs |
|
|
FRT P |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
5’ |
3’ |
|
|
|
|
|
|
|
|
|
|
|
|
RS5 (w+) |
|
|
|
|||
HindIII |
|
|
|
|
|
HindIII HindIII |
|
|
|
|
|
|
||||
0 |
1.0 |
2.0 |
3.0 |
4.0 |
5.0 |
6.0 |
7.0 |
|
8.0 |
|
|
|
|
FLP |
|
|
Конструкции, содержащие FLP è FRT äëÿ |
|
|
|
|
|
|
||||||||||
трансформации дрозофилы. а – вектор |
RS5r (w) |
|
|
|
||||||||||||
P[ry+,hsFLP], á – P[FRTwhsFRT] (Èç: Golic, |
|
|
|
|
|
|
||||||||||
Lindquist, 1989). |
|
|
|
|
|
|
|
|
|
|
|
|
||||
как и лямбда-интеграза, каждый из этих |
á |
|
|
5’ |
3’ |
|
||||||||||
ферментов |
опознает |
специфические |
|
|
|
|
||||||||||
RS3 (w+) |
|
|
|
|||||||||||||
последовательностиДНКвсоответствующем |
|
|
|
|||||||||||||
мобильном элементе, чья рекомбинация |
|
|
|
|
FLP |
|
||||||||||
катализируется.Вотличиеотинтегразылямбда |
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|||||||||||
фага,этиинтегразынетребуютспецифических |
|
|
|
|
|
|
||||||||||
последовательностей ДНК в хромосоме |
RS3r (w) |
|
|
|
||||||||||||
мишени и они не формируют сочленения |
|
|
|
|
|
|
||||||||||
гетеродуплекса. Вместо этого они образуют |
â |
a |
|
|
b |
c |
||||||||||
надрезы с обоих концов линейной |
|
|
|
|
|
|
||||||||||
последовательности мобильного элемента и |
|
f |
|
|
e |
|
||||||||||
затемкатализируютпрямуюатакуэтихконцов |
|
|
|
d |
||||||||||||
|
|
|
|
|
||||||||||||
ДНК на мишень ДНК, разрывая в ней |
|
|
|
|
|
|
||||||||||
фосфодиэфирные связи. В результате в |
|
|
|
|
|
|
||||||||||
рекомбинантной молекуле ДНК образуются |
|
a |
5’ |
3’ |
e |
|
||||||||||
две коротких однонитчатых бреши, по одной |
|
|
||||||||||||||
|
|
|
|
|
|
|||||||||||
на каждом конце мобильного элемента. Они |
|
|
|
|
|
d |
||||||||||
заполняются ДНК-полимеразой для |
|
|
|
w+ |
|
|
||||||||||
завершения процесса рекомбинации (Рис. |
|
|
|
|
|
c |
||||||||||
8.28). В соответствии с этим механизмом |
|
f |
|
|
b |
|||||||||||
|
|
|
|
|||||||||||||
образуются короткие дупликации в ДНК |
Получение инверсии с помощью двух |
|||||||||||||||
клетки-хозяинеприлежащей кместуинсерции. |
||||||||||||||||
противоположно ориентированных FRT |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||
Такие фланкирующие дупликации являются |
районов. а, б - получение линий с RS5 (а) и |
|||||||||||||||
отличительной чертой случайной или |
RS3r (б) элементами в хромосомах; в – |
|||||||||||||||
транспозиционной сайт-специфичной |
использование этих элементов для получения |
|||||||||||||||
рекомбинации. |
|
|
|
|
|
|
|
инверсии; · – черный кружок – центромера. |
||||||||
|
Фермент-интеграза этого типа впервые |
Затемненные участки – части гена white, |
||||||||||||||
впервые была выделена в активной форме из |
мРНК – нарисована над геном, черные |
|||||||||||||||
фактериофагаMu. |
|
|
|
|
|
|
полустрелки – FRT, маленькие стрелки – |
|||||||||
|
|
|
|
|
|
|
|
|
|
концевые участки P-элемента. (w+) èëè (w) – |
||||||
|
|
|
|
|
|
|
|
|
|
получаемые фенотипы мух (Из: Golic, Golic, |
||||||
|
|
|
|
|
|
|
|
|
|
1996, P. 1695). |
|
|
|
|||
224

Молекулярные механизмы мутагенеза... |
|
|
|
Глава 8 |
||
|
|
|
|
|
||
Сингер М., Берг П. Гены и геномы. Москва, |
8.4. Генная конверсия |
|
||||
Ìèð, ò. 1, 103-111, 1998. |
|
Иногда в результате мейоза получается |
||||
Смирнов В.Г. Цитогенетика. Москва, Высшая |
||||||
трикопииматеринскогоаллеля и толькоодна |
||||||
школа, 42-107, 1991. |
|
копия отцовского, что свидетельствует об |
||||
Alberts B., Bray D., Lewis J., Raff M., Roberts K., |
||||||
Watson J.D. Molecular biology of the cell |
изменении одной копии отцовского аллеля в |
|||||
(Third edition). Garland Publishing Inc. New |
материнскую. Это явление называется |
|||||
York, London, 264-268, 1994. |
генной конверсией. Оно часто происходит в |
|||||
Carpenter A.T.C. Gene conversion, recombination |
связи с событиями общей рекомбинации и |
|||||
nodules, and the initiation of meotic synapsis. |
репарацииДНК. |
|
||||
BioEssays 6: 232-236, 1988. |
Как уже отмечалось выше, в ходе |
|||||
Golic K.G., Golic M.M. Engeneering the Drosophila |
мейоза |
образуется |
сочленение |
|||
genome: chromosome rearrangements by |
гетеродуплекса в участках кроссинговера |
|||||
design. Genetics 144: 1693-1711, 1996. |
между гомологичными материнской и |
|||||
Golic K.G., Lindquist S. The FLP recombinase of |
||||||
отцовской хромосомами. Если эти участки |
||||||
yeast catalyzes site-specific recombination in |
хромосом несколько различаются, в участке |
|||||
the Drosophila genome. Cell 59: 499-509, |
||||||
1989. |
|
сочленения могут произойти нарушения |
||||
|
спаривания нуклеотидов (mismatch). Ýòè |
|||||
Klein H.L. Genetic control of intrachromosomal |
||||||
recombination. BioEssays 17: 147-159, 1995. |
нарушения будут исправляться системой |
|||||
Lewin B. Genes, Fifth edition. Oxford University |
репарации ДНК, в результате чего либо будут |
|||||
Press. Oxford, New York, Tokyo, 967-975, |
удалены нуклеотиды на отцовской молекуле |
|||||
1994. |
|
или наоборот. Следствием этого нарушения |
||||
Sybenga J. Recombination and chiasmata: few but |
спаривания будет генная конверсия. Генная |
|||||
intriguing discrepancies. Genome 39: 473-484, |
конверсия может также произойти по ряду |
|||||
1996. |
|
других механизмов, но все они требуют |
||||
|
|
|||||
|
|
осуществление какого-то варианта событий |
||||
|
|
общей рекомбинации, по которому две |
||||
|
|
|
|
|
|
|
Рисунок 8.28 |
|
|
|
|
|
|
ДНК вируса |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Интеграза |
|
|
|
|
|
|
|
|
|
|
|
надрезает |
|
Интеграза |
|
|
|
|
|
|
|
|
|
концы ДНК вируса |
|
|||
|
|
|
|
|
|
|
|
|
|||
|
Хромосома хозяина |
5’ |
|
|
|
|
OH 3’ |
HO |
OH |
||
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|||||
|
3’ HO |
|
|
|
|
5’ |
|
|
|||
|
|
|
|
|
|
||||||
5’ |
|
|
3’ |
|
|
|
|||||
|
|
|
|
Присоединение вирусной ДНК |
|
||||||
3’ |
|
|
5’ |
|
|
к хромосоме хозяина |
|
|
|||
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
|
|
|
|
|
|
|
|
|
|
|||
3’ |
|
|
|
|
|
|
|
|
|
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
||||||||||||
|
|
Зашивание пробелов в ДНК |
|||||||||||
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
Встроенная ДНК вируса |
|||||||
Хромосома хозяина 5’ |
|
|
|
|
|
|
|
|
|
|
|
|
3’ |
|
|
|
|
|
|
|
|
|
|
|
|
||
3’ |
|
|
|
|
|
|
|
|
|
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
||
Короткие прямые повторы хромосомной ДНК
Схема случайной рекомбинации (Из: Alberts et al., 1994, P. 273).
225
Глава 8 |
...Молекулярные механизмы мутагенеза |
|
|
гомологичные молекулы ДНК располагаются вместе. Поскольку производятся дополнительные копии фрагментов ДНК, процесс конверсии происходит с ограниченным синтезом ДНК. Опыт показывает, что обычно только малые участки ДНК испытывают генную конверсию и в большинстве случаев изменяется только часть гена.
Генная конверсия может происходить в митотических клетках, но значительно реже. Один из весьма вероятных механизмов генной конверсии как в мейотических, так и в митотических клетках показан на Рис. 8.29.
Литература к разделу 8.4.
Глазер В.М. Конверсия гена. Соросовский образовательный журнал $: 23-31, 2000.
Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J.D. Molecular biology of the cell (Third edition). Garland Publishing Inc. New York, London, 268-270, 1994.
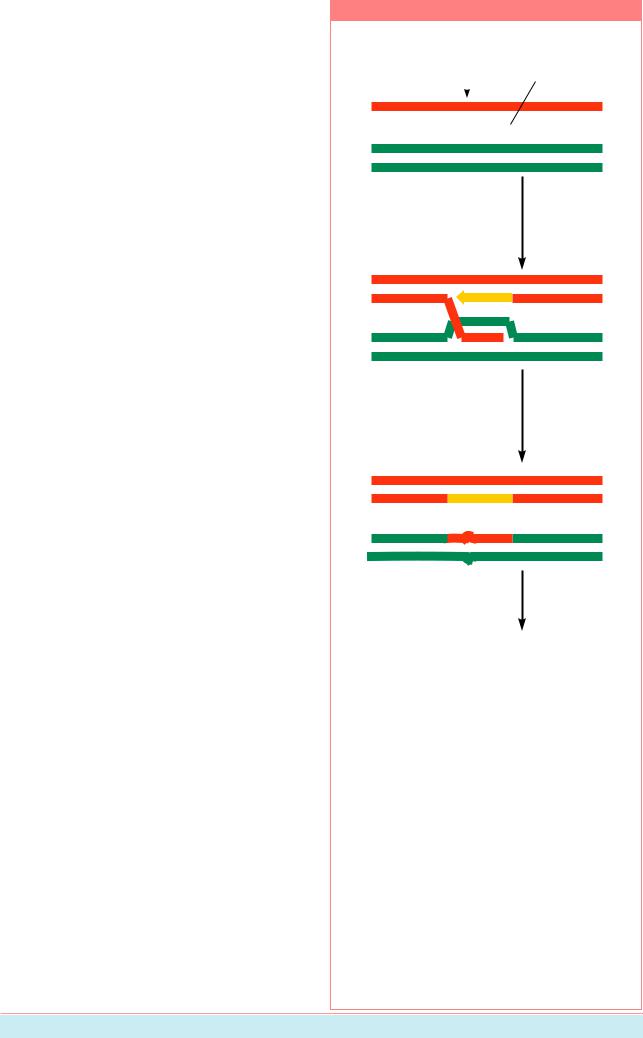
Рисунок 8.29
ñàéò â ãåíå Õ, ïî |
|
|||
которому красный |
одноцепочечный |
|||
и зеленый аллели |
||||
разрыв |
||||
различаются |
|
|
||
|
|
|
||
|
|
|||
|
|
|
|
|
5’ |
......................................... |
3’ |
|||
3’ |
|
|
|
5’ |
|
|
|
|
|||
5’ 3’ |
|||||
|
|
||||
.........................................
ДНК-полимераза перемещает одну цепь красной спирали, которая спаривается с зеленой спиралью
.............. .........................
.............. ........ ................
ДНК-полимераза останавливается, излишек одноцепочечной ДНК разрушается нуклеазами, все оставшиеся разрывы зашиваются лигазами
.........................................
................ .......................
Репликация ДНК образует три красных аллеля и один зеленый аллель гена Õ
Один зеленый аллель был конвертирован в красный
Один из механизмов общей рекомбинации может приводить к генной конверсии. Процесс начинается с образования ника на одной из цепей ДНК (красная на рисунке). На первом этапе ДНК-полимераза начинает синтез дополнительной копии цепи в красной спирали, заменяя оригинальную копию. Эта одиночная цепь затем спаривается с гомологичным районом зеленой цепи. На этапе 2 короткий район неспаренной зеленой цепи, продуцированной на этапе 1, деградирует, завершая перенос нуклеотидных последовательностей. Результат обычно виден в следующем клеточном цикле, после того как репликация ДНК разделяет две некомплементирующие цепи (Из: Alberts et al., 1994, p. 269).
226