

Липопротеины − транспортные формы липидов
|
|
Хиломикро- ны |
ЛПОНП |
ЛППП |
ЛПНП |
ЛПВП |
|
Состав, % Белки ФЛ ХС ЭХС ТАГ |
2 3 2 3 85 |
10 18 7 10 55 |
11 23 8 30 26 |
22 21 8 42 7 |
50 27 4 16 3 |
|
Функции |
Транспорт липидов из клеток кишечника (экзогенных липидов) |
Транспорт липидов, синтезируемых в печени (эндогенных липидов) |
Промежуточ- ная форма превращения ЛПОНП в ЛПНП под действием фермента ЛП-липазы |
Транспорт холестерола в ткани |
Удаление избытка холестерола из клеток и других липопротеинов. Донор апопротеинов А, С-II |
|
Место образования |
Эпителий тонкого кишечника |
Клетки печени |
Кровь |
Кровь (из ЛПОНП и ЛППП) |
Клетки печени − ЛПВП-пред- шественники |
|
Плотность, г/мл |
0,92−0,98 |
0,96−1,00 |
|
1,00−1,05 |
1,06−1,21 |
|
Диаметр частиц, нМ |
Больше 120 |
30−100 |
23−30 |
20−25 |
7−15 |
|
Основные аполипопротеины |
В-48, С-II, Е |
В-100, Е (С-II) |
В-100, Е (С-II) |
В-100 |
А-I, А-II, С-II, Е |
Примечание: ФЛ фосфолипиды, ХС холестерин, ЭХС эфиры холестерина, ТАГ триацилглицериды. Апопротеины: В-48 основной белок ХМ, В-100 основной белок ЛПОНП, ЛПНП, ЛППП, взаимодействует с рецепторами ЛПНП; С-II активатор липопротеинлипазы (ЛП-липазы), переносится с ЛПВП на ХМ и ЛПОНП в крови; Е участвует в связывании липопротеинов с рецепторами ЛПНП и другими рецепторами, А-I активатор лецитинхолестеринацилтрансферазы (ЛХАТ) (согласно [10])
Липопротеины отличаются по своей функции.
1. Хиломикроны (ХМ) образуются в клетках кишечника, их функция: перенос экзогенного жира из кишечника в ткани (в основном в жировую ткань), а также транспорт экзогенного холестерина из кишечника в печень.
2. Липопротеины очень низкой плотности (ЛПОНП) образуются в печени, их роль: транспорт эндогенного жира, синтезированного в печени из углеводов, в жировую ткань.
3. Липопротеины низкой плотности (ЛПНП) образуются в кровеносном русле из ЛОНП через стадию образования липопротеинов промежуточной плотности (ЛППП). Их роль: транспорт эндогенного холестерина в ткани.
4. Липопротеины высокой плотности (ЛПВП) образуются в печени, основная роль транспорт холестерина из тканей в печень, т. е. удаление холестерина из тканей, а далее холестерин выводится с желчью.
5.3.8.1. Структура липопротеинов. Независимо от типа все липопротеины имеют сходное строение.
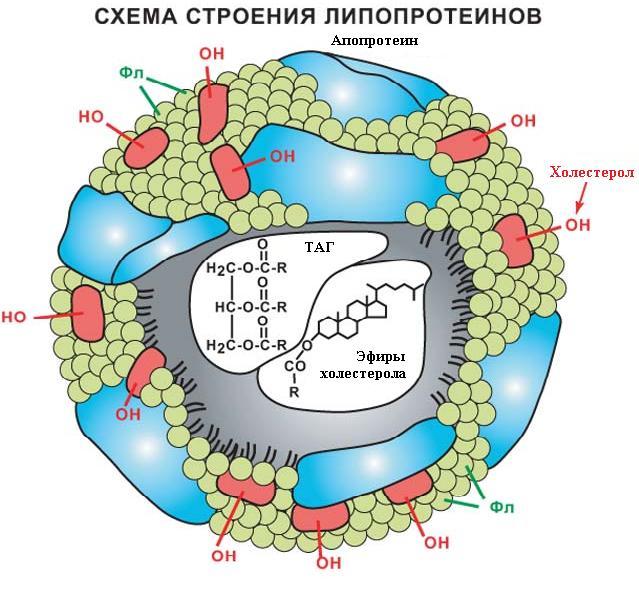
Они представляют собой сферические частицы, в которых можно выделить гидрофобную сердцевину, состоящую из ТАГ и эфиров холестерина (ЭХС) и гидрофильную оболочку, в составе которой – фосфолипиды, гликолипиды и белки. Компоненты частиц связаны слабыми типами связей и находятся в состоянии постоянной диффузии – способны перемещаться друг относительно друга. Белки, входящие в состав липопротеина и называемые апопротеинами (обозначаются латинскими буквами), могут быть или интегральными, не способными к отделению от липопротеина, т. е. присущи только этому типу липопротеина, или свободно переносимыми от одного типа липорпотеина к другому типу.
Функции апопротеинов в составе липопротеинов заключаются в: 1) формировании структуры липопротеинов; 2) взаимодействии с рецепторами на клеточной поверхности, тем самым определяется, с какими тканями связывается данный тип липопротеина; 3) активации ферментов липидного обмена. Иногда апопротеины сами обладают собственной ферментативной активностью; 4) выполнении функции эмульгаторов, так как апопротеины являются гидрофильными веществами; 5) транспорте липидов от одного типа липопротеина к другому.
5.3.8.2. Хиломикроны. Из ресинтезированных ТАГ, эфиров холестерина, фосфолипидов, поступивших с пищей жирорастворимых витаминов образуются комплексы липопротеинов, получившие название хиломикроны (ХМ), функция которых заключается в доставке экзогенных (пищевых) жиров в периферические ткани. ХМ содержат около 2 % белка, 7 % фосфолипидов, 8 % холестерина и его эфиров и более 80 % ТАГ. Диаметр ХМ колеблется от 0,1 до 5 мкм. Из-за больших размеров частиц ХМ не способны проникать из эндотелиальных клеток кишечника в кровеносные капилляры и диффундируют в лимфатическую систему кишечника, а из нее – в грудной лимфатический проток, из которого ХМ попадают в кровяное русло, где осуществляют транспорт ТАГ, холестерина и частично фосфолипидов из кишечника через лимфатическую систему в кровь.
Через 1–2 ч после приема пищи, содержащей жиры, повышается концентрация ТАГ в крови и появляются в кровеносном русле ХМ. Через 10–12 ч после приема пищи содержание ТАГ возвращается к нормальным величинам, а ХМ полностью исчезают из крови.
ХМ свободно диффундируют из плазмы крови в межклеточные пространства печени. Гидролиз ТАГ, содержащихся в ХМ, происходит как внутри печеночных клеток, так и на поверхности. ХМ не способны (из-за своих размеров) проникать в клетки жировой ткани, поэтому ТАГ ХМ подвергаются гидролизу на поверхности эндотелия капилляров жировой ткани при участии фермента липопротеинлипазы.
Основной апопротеин в составе ХМ белок апоВ-48. Белок кодируется тем же геном, что и белок В-100, входящий в состав ЛПОНП, ЛПНП, ЛППП (см. таблицу) и синтезируемый в печени. В кишечнике происходит считывание лишь части гена, а именно 48 %, поэтому белок и получил свое название В-48, синтезируется он в шероховатом эндоплазматическом ретикулуме и гликозилируется. В аппарате Гольджи затем формируются так называемые «незрелые» ХМ. Они путем экзоцитоза попадают в главный грудной лимфатический проток, а через него в кровь. В лимфе и крови ХМ получают апопротеины Е и С-II, превращаясь в «зрелые» ХМ. После приема жирной пищи образовавшиеся ХМ опалесцируют, придавая плазме крови похожий на молоко вид. Транспортируя липиды к различным тканям, где они подвергаются расщеплению, ХМ постепенно исчезают из крови и плазма становится прозрачной.