

Общая генетика / glava4
.pdf
|
|
|
Глава 4. Изменчивость наследственного материала |
||||||
активность гена не проявляется и 0- |
чувствительные (180C) ts (temperature |
|
|||||||
антиген не |
модифицируется. Это |
sensitive) мутации и теплочувствительные |
|||||||
обстоятельство объясняет почему А и В |
(290C) ts-мутации. При этом температура, |
||||||||
аллели доминируют у А0 и В0 гетерозигот: |
при которой сохраняется нормальный |
||||||||
соответствующая |
трансферазная |
фенотип, составляет 250C. |
|
|
|||||
активность (À èëè Â) создает À èëè Â |
Мутации, |
чувствительные к |
|||||||
антиген. А и В аллели кодоминируют в |
стрессу. В данном случае мутанты |
|
|||||||
гетерозиготе |
|
ÀÂ, |
поскольку |
развиваются нормально, если их не |
|||||
экспрессируются активности обеих |
подвергнуть каким-либо стрессирующим |
||||||||
трансфераз. Гомозигота 00 является нуль- |
воздействиям. Так, мутанты sesB (stress- |
||||||||
аллелем, и поэтому не имеет ни одного |
sensitive) у дрозофилы не проявляют |
||||||||
антигена. Таким образом, на базе одного |
каких-либо отклонений. Если резко |
|
|||||||
гена получается три аллеля (Из: Lewin, |
встряхнуть пробирку, у мух начинаются |
||||||||
1994, p. 77). |
|
|
|
судороги и они неспособны двигаться. |
|||||
4.1.8. Условные мутации |
|
Ауксотрофные мутации. Обычно |
|||||||
|
бактерий высевают на чашки Петри, |
||||||||
В ряде случаев мутантный фенотип |
|||||||||
содержащие полную среду, в состав |
|||||||||
возникшей мутации становится видимым |
которой |
входят все |
необходимые |
||||||
только при выполнении определенных |
питательные вещества, нужные для роста. |
||||||||
условий. Этими условиями могут быть |
Есть еще минимальная среда, состоящая |
||||||||
изменения температуры или условий |
из агара, воды, сахара и солей. |
|
|||||||
существования. |
|
|
Нормальные бактерии способны сами |
||||||
Температурно-чувствительные |
синтезировать необходимые им сложные |
||||||||
мутации (см. Suzuki et al., 1976, p. 208). |
органические соединения (витамины, |
||||||||
Мутанты этого типа живут и развиваются |
аминокислоты, нуклеотиды) и могут жить |
||||||||
нормально при одной (пермиссивной) |
на минимальной среде, а мутантные - не |
||||||||
температуре и проявляют отклонения при |
могут. |
Таких |
мутантов называют |
||||||
другой (рестриктивной). Например, |
ауксотрофными. Они могут выживать |
||||||||
мутация shi у дрозофилы. При 250C |
только на полной или же на минимальной, |
||||||||
мутанты не проявляют каких-либо |
но с добавкой нормального продукта того |
||||||||
отклонений, при 290С - у них наступает |
гена, который в данной линии мутировал. |
||||||||
полный паралич. Полагают, что в |
4.2. Спонтанные и |
|
|
||||||
результате мутации происходит замена |
|
|
|||||||
аминокислоты в молекуле белка, однако |
индуцированные мутации |
|
|||||||
при одной температуре влияние этой |
4.2.1. Методы учета мутаций |
|
|||||||
мутации на конформацию молекулы не |
Для учета частот возникновения или |
||||||||
сказывается. При другой температуре |
|||||||||
выявления мутаций |
используют |
||||||||
конформация меняется, и тут оказывается, |
|||||||||
что в белке произошло нарушение |
различные методические приемы. Первые |
|
|||||||
структуры. |
Выделяют |
холодо- |
методы были предложены Г. Меллером |
|
|||||
4-11
Глава 4. Изменчивость наследственного материала
|
ClB |
|
îïûò |
|
À P |
|
|
|
|
|
+ + + |
|
|
|
|
|
ClB |
+ + + |
|
F1 |
|
îïûò |
|
|
|
ClB |
îïûò |
ClB |
îïûò |
F2 |
+ + + |
+ + + |
|
|
|
|
|
2: 1
ClB |
|
îïûò lm |
|
Á P |
|
|
|
+ + + |
|
|
|
|
ClB |
+ + + |
|
F1 |
îïûò lm |
|
|
|
|
|
|
ClB |
îïûò lm |
ClB |
îïûò lm |
F2 |
|
|
|
+ + + |
+ + + |
|
|
2: 0
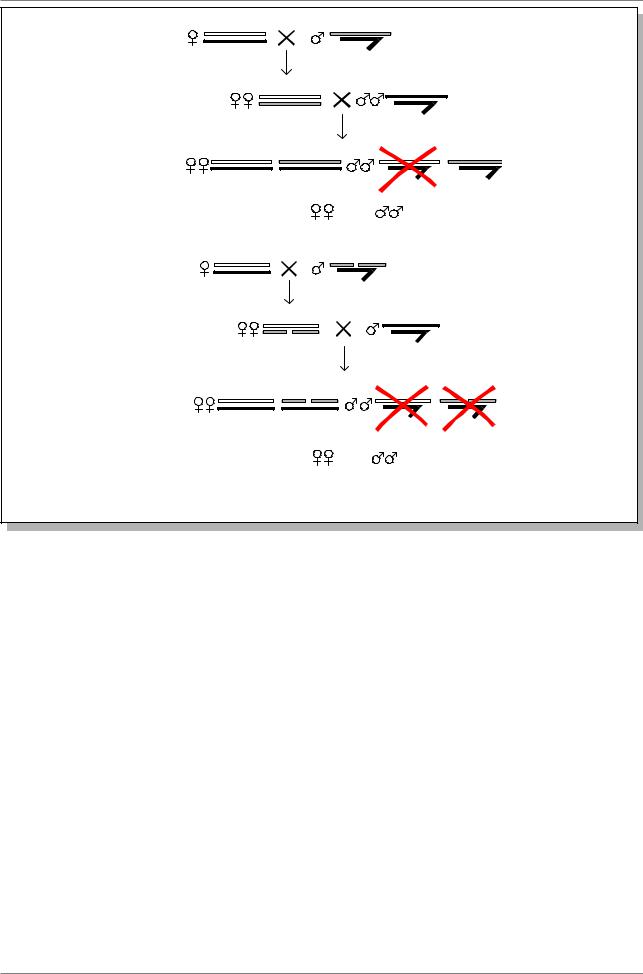
Рис. 4.5. Схема скрещиваний для выявления летальных мутаций в X- хромосоме дрозофилы
для определения частот образования мутаций у дрозофилы.
Метод ClB. Наиболее объективно можно учитывать частоту возникновения рецессивных летальных мутаций, приводяших в гомозиготном состоянии к смерти несущих их особей. Генетическая структура ClB характеризуется тем, что одна из Х-хромосом маркирована доминантным геном Bar (B) и инверсией, названной С. Эта инверсия препятствует кроссинговеру и обладает рецессивным летальным эффектом - l. По этому линия и названа - ClB. Самок из этой линиианализатора скрещивают с самцами из анализируемой выборки. Эти самцы могут быть взяты из природной популяции и тогда можно оценить
частоту леталей данной популяции. Или же самцов обрабатывали мутагеном. В этом случае оценивается частота летальных мутаций, вызванных этим мутагеном. В F1 отбирают самок ClB/+, гетерозиготных по мутации Bar, и скрещивают индивидуально (в отдельной пробирке) с самцом. Если в опытной хромосоме нет мутации, то в потомстве будет два класса самок и один класс самцов (B+), поскольку самцы ClB гибнут из-за наличия летали l, т.е. общее расщепление по полу будет 2:1 (Рис. 4.5. А).
Если же в опытной хромосоме есть летальная мутация lm, то после скрещиваний аналогичных выше описанным, произойдет изменение в
4-12

Глава 4. Изменчивость наследственного материала
À
Á
P
F1
F2
F3
P
F1
F2
F3
Cy |
L |
îïûò |
|
|
|
Pm |
|
îïûò |
|
|
|
|
|
|
|
|
|
|
Cy L |
|
|
Cy L |
|
|
Pm |
|
|
îïûò |
|
|
Cy L |
|
Cy L |
|
|
|
îïûò |
|
îïûò |
|
|
Cy |
L |
Cy L |
Cy |
L |
îïûò |
îïûò |
îïûò |
Cy |
L |
îïûò |
|
|
|
|
|
||
|
2 Cy L : |
1 Cy+ L+ |
|
||
Cy |
L |
îïûò lm |
|
|
|
Pm |
îïûò |
|
|
|
|
|
|
|
|
||
|
Cy |
L |
|
Cy |
L |
|
îïûò lm |
|
Pm |
||
|
Cy |
L |
|
Cy |
L |
|
îïûò lm |
|
îïûò lm |
||
Cy |
L |
Cy L |
Cy |
L |
îïûò lm |
îïûò lm |
îïûò lm |
Cy |
L |
îïûò lm |
|
|
|
2 Cy L |
: 0 Cy+ L+ |
||
Рис. 4.6. Схема скрещиваний для выявления летальных мутаций во второй хромосоме дрозофилы
расщеплении по полу в F2. Â F2 гибнут самцы обоих классов в одном случае изза наличия летали в Х-хромосоме ClB, в другом из-за наличия летали lm в опытной Х-хромосоме (Рис. 4.5. Б). Определяя отношение числа Х-хромосом (или пробирок с индивидуальными скрещиваниями), в которых возникла леталь, к общему числу изученных Х- хромосом (пробирок), подсчитывают частоты нахождения леталей в
определенной группе или выборке дрозофилы.
Меллер неоднократно модифицировал свой метод выявления леталей в Х-хромосоме дрозофилы, в результате чего появились такие линиианализаторы, как Mu-5, Basc, Binsn.
Для учета летальных мутаций в аутосомах дрозофилы используют линии сбалансированных леталей. Рассмотрим метод обнаружения леталей во второй хромосоме: метод Cy L/Pm.
4-13

Глава 4. Изменчивость наследственного материала
Âэтой линии в одной хромосоме расположены доминантные мутации Cy (Curly - загнутые крылья) и L (Lobe - маленькие дольковидные глаза), каждый из которых в гомозиготном состоянии вызывает летальный эффект. Мутации сопряжены с инверсией. Во второй хромосоме, также несущей инверсию, расположена доминантная мутация Pm (Plum - коричневые глаза). Испытываемого самца скрещивают с самкой из линии Cy L/Pm (на схеме показаны не все классы потомков).
ÂF1 отбирают самцов Cy L/+. У них подавлен кроссинговер. Самцов индивидуально скрещивают с самками
исходной линии Cy L/Pm. В F2 отбирают самцов и самок Cy L, у которых вторая хромосома является испытуемой. В результате скрещивания их между собой получается 4 класса потомков. Один из них погибает из-за гомозиготности по мутациям Cy и L, еще два класса потомков это гетерозиготы Cy L/+ и один класс - это гомозиготы по испытуемой хромосоме. В итоге получается 2 класса мух Cy L и один класс Cy+ L+ (Рис. 4.6. А). Если в испытуемой хромосоме произошла летальная мутация, в потомстве от последнего скрещивания будут только мухи Cy L (Рис. 4.6. Б). С помощью такого метода можно учитывать частоту рецессивных летальных мутаций во второй хромосоме дрозофилы.
Учет мутаций у микроорганизмов
(см. Инге-Вечтомов, 1989, стр. 298). Использование микроорганизов очень удобно из-за того, что они имеют гаплоидный набор, и мутации проявляются уже в первом поколении.
Кроме того, каждая клетка на плотной среде может образовать отдельную колонию, представляющую собой клон идентичных клеток.
Если получают мутации, дающие селективное преимущество, то мутантов легко выявить методом отпечатков или реплик, предложенным Э. и Дж. Ледербергами (рис. 4.7.).
При изучении мутаций устойчивости E. coli к бактериофагу Т1 клетки бактерий (мутанты TonS) высевают на агаризованную среду в чашки Петри таким образом, чтобы на них образовались отдельные колонии. Затем при помощи бархатной печатки эти колонии перепечатывают на чашки с нанесенной суспензией частиц фага Т1. Большая часть клеток исходной чувствительной (TonS) культуры не будет образовывать колоний, поскольку их лизирует бактериофаг. Вырастут лишь отдельные мутантные колонии (Tonr), устойчивые к фагу. Подсчитывая число колоний в контрольном и опытном
Рис. 4.7. Метод отпечатков для обнаружения мутантов у бактерий устойчивых к фагу T1: 1 - получение отпечатка колоний на бархате, 2 - перепечатка на среду, 3 - инкубация отпечатка. Растущие колонии - черные
4-14

Глава 4. Изменчивость наследственного материала
(например, после облучения ультрафиолетовым светом) вариантах, легко определить частоту индуцированных мутаций.
4.2.2.Спонтанные мутации
Âлюбой популяции живых организмов, живущих на Земле, всегда есть особи, несущие мутации. Многие годы до открытия возможности искуственной индукции мутаций селекционеры и исследователи наследственности, включая Менделя и Моргана, использовали мутации этого типа. Их называют спонтанными.
Начиная с 1925 года С.С. Четвериков и его молодые коллеги Б.Л. Астауров, Н.К. Беляев, С.М. Гершензон, П.Ф. Рокицкий, Д.Д.Ромашовврезультате экспериментальной проверки природных популяций дрозофилы нашли в них большое число различных мутаций. Каждый ген с той или иной частотой спонтанно переходит в мутантное состояние (табл. 4.5.).
[Есть данные о частотах спонтанных мутацийвучебникеИнге-Вечтомова,1989, с. 302-303].
Причины индукции спонтанных мутаций не ясны. Долгое время полагали, что к числу индуцирующих факторов относится естественный фон ионизирующих облучений, образуемый доходящими до поверхности земли космическими лучами, гаммаизлучениями Земли и радиоактивными веществами (40Ê, 14С, Rn), поступающими в малых
количествах в организм из окружающей среды. Однако, как показали расчеты, у дрозофилы естественый радиационный
фон может быть ответствен только приблизительно за 0,1% спонтанных мутаций. Хотя, по мере увеличения продолжительности жизни организма воздействие естественного фона может накапливаться, и у человека от 1/4 до 1/10 спонтанных мутаций может быть отнесено за счет естественного фона радиации (из: Гершензон, 1983, с. 237).
Второй причиной спонтанных мутаций являются случайные повреждения хромосом и генов в ходе нормальных метаболических процессов, происходящих в клетке. По многочисленным данным спонтанные мутации возникают во время деления хромосом и репликации ДНК. Считают вероятным, что спонтанные мутации представляют чаще всего следствие случайных ошибок в функционировании молекулярных механизмов.
Третьей причиной спонтанных мутаций является перемещение по геному мобильных элементов, (см. раздел 6.6.), которые могут внедриться в любой ген и вызвать в нем мутацию. По расчетам американского генетика Мелвина Грина около 80% спонтанных мутаций приходится на счет перемещений мобильных элементов.
4.2.3. Индуцированные мутации
История попыток индукции мутаций знает много примеров. Тут и неудачная попытка Т.Х. Моргана и весьма удачная попытка Надсона и Филиппова, в СССР,
которые, облучая рентгеновскими лучами культуры плесневых грибов, Mucor genevensis, в 1925 году получили расщепление облучаемой культуры “на
4-15

Глава 4. Изменчивость наследственного материала
Табл. 4.5. Частота спонтанных мутаций некоторых генов (из: Гершензон, 1983, с. 222)
Тип, класс |
Âèä |
Мутационное |
Направление |
Частота мутаций |
|
|
изменение |
мутирования |
|
Млекопита- |
Человек |
Альбинизм |
+ → a |
2,8 - 3,3×10-5 |
þùèå |
|
Фенилкетонурия |
+ → ph |
2,5 - 8×10-5 |
|
|
Микроцефалия |
+ → mc |
2,7×10-5 |
|
|
Гемофилия |
+ → h |
2 - 3,2×10-5 |
|
|
Аниридия |
+ → Anir |
0,5×10-5 |
|
Ìûøü |
Ослабленная окраска |
+ → d |
3×10-5 |
|
|
Альбинизм |
+ → c |
3×10-5 |
|
|
Пегость |
+ → s |
3×10-5 |
Насекомое |
Дрозофила |
Желтое тело |
+ → y |
1×10-5 (у самок) |
|
|
Òî æå |
+ → y |
1×10-4 (у самцов) |
|
|
Белые глаза |
+ → w |
2-4×10-5 |
|
|
Вильчатые щетинки |
+ → f |
2,9×10-5 |
|
|
Òî æå |
f → + |
1,5×10-5 |
|
|
Вырезки на крыльях |
+ → ct |
1,5×10-4 |
|
|
Коричневые глаза |
+ → bw |
3×10-5 |
Цветковое |
Кукуруза |
Пурпурный эндосперм |
+ → pr |
1,1×10-5 |
растение |
|
Сахарный эндосперм |
+ → su |
2,4×10-6 |
|
|
Морщинистый |
+ → sh |
1,2×10-6 |
|
|
эндосперм |
||
Водоросль |
Хламидомонада Устойчивость к стреп- |
+ → strr |
1×10-6 |
|
|
|
томицину |
||
Грибы |
Нейроспора |
Потребность в аденине |
ade- → ade+ |
4×10-8 |
|
|
Потребность в инозитоле |
ino- → ino+ |
2-8×10-8 |
|
Пекарские |
Потребность в метионине |
met- → met+ |
3,4 - 6,5×10-8 |
|
дрожжи |
|
|
|
Бактерии |
Кишечная |
Потребность в гистидине |
his+ → his- |
2×10-8 |
|
палочка |
Òî æå |
his- → his+ |
2×10-6 |
|
|
Устойчивость к стреп- |
str-s → str-d |
2×10-8 |
|
|
томицину |
lac- → lac+ |
1×10-9 |
|
|
Потребность в лактозе |
2×10-7 |
|
|
|
Устойчивость к фагу Т5 |
T5s → T5r |
7×10-8; 1 |
|
Золотистый |
Устойчивость к сульфа- |
sul- → sul+ |
1×10-9 |
|
стафиллококк |
ìèäó |
|
|
Вирусы |
Ôàã Ò2 |
Изменение круга хозяев |
h+ → h- |
3×10-9 |
|
Ôàã Ò4 |
Òî æå |
+ → rIII |
2×10-5 |
|
|
|
+ → auc |
1×10-7 |
|
Вирус |
Мозаичность типа аукуба |
1,6×10-8 |
|
мозаичности
табака
Примечание. Частота мутаций указанных в таблице генов приведена для вирусов на один цикл размножения, для бактерий и дрожжей - на одно клеточное деление, для прочих организмов - на одно поколение.
две формы или расы”. “Таким образом получились две формы, два мутанта, отличающиеся не только друг от друга, но и от исходной (нормальной) формы”. Мутанты оказались стабильными, т.к. “дальнейшие восемь генераций (восемь последовательных пересевов) уже не
подвергались действию рентгеновских лучей и тем не менее сохраняли приобретенные свойства: они оказались стойко наследственно закрепленными” (см. статью Надсона и Филиппова, 1925, в сборнике “Классики советской генетики”, стр. 122). Эта статья была
4-16

Глава 4. Изменчивость наследственного материала
Табл. 4.6. Индукция видимых мутаций экзогенной ДНК у D. melanogaster (Из: Gershenson, Alexandrov, 1997, p. 186).
|
Эксперимент |
|
Контроль |
|
||||
|
|
|
|
|
|
|
|
|
|
1939 ã. |
1940 ã. |
Всего |
1939 ã. |
|
1940 ã. |
|
Всего |
|
|
|
|
|
|
|
|
|
Число скрещиваний |
228 |
422 |
650 |
208 |
|
413 |
|
621 |
|
|
|
|
|
|
|
|
|
Число мух F1 |
12684 |
20761 |
33445 |
14481 |
|
23401 |
|
37882 |
|
|
|
|
|
|
|
|
|
Число мутаций |
13 |
25 |
38 |
0 |
|
0 |
|
0 |
|
|
|
|
|
|
|
|
|
опубликована на русском языке, к тому же
âработе не использовались какие-либо методы количественной оценки действия рентгеновских лучей, и в целом она осталась малозамеченной.
Â1927 году Г.Дж. Меллер сообщил о действии рентгеновских лучей на мутационный процесс у дрозофилы и предложил ставший классическим количественный метод учета рецессивных летальных мутаций в Х-хромосоме (см. Рис. 4.6.).
[Чуть позже, в 1928 году Л. Стадлер
âСША описал влияние рентгеновских лучей на мутационный процесс у ячменя (проверять).
Сапегин - ? - кукуруза - 1929 г. (проверять)].
Â1930-õ годах был открыт химический мутагенез у дрозофилы: В.В. Сахаров (1932), М.Е. Лобашев и Ф.А. Смирнов (1934) показали, что некоторые соединения, такие как йод, уксусная кислота, аммиак, способны
Дополнение4.1.
В 1946 году за открытие радиационного мутагенеза Г.Дж.Меллеру была присуждена Нобелевская премия.
индуцировать рецессивные летали в Х- хромосоме. В 1939 году С.М. Гершензон открыл сильный мутагенный эффект экзогенной ДНК у дрозофилы.
Под влиянием идей Н.К. Кольцова о том, что генная нить в хромосомах является цепью больших органических молекул или возможно одиночной гигантской молекулой, С.М. Гершензон решил проверить свое предположение, что именно ДНК является такой молекулой. Он изолировал ДНК из тимуса и добавил ее в корм личинкам дрозофилы. Среди 15 тысяч проанализированных контрольных мух (т.е. без ДНК в корме) не было получено ни одной мутации, в то время как в опыте среди примерно 13 тыс. мух были получены 13 видимых мутаций (Табл. 4.6.).
В 1941 году Ш. Ауэрбах и Дж.М. Робсон, используя метод ClB Меллера, показали, что горчичный газ (азотистый иприт) индуцирует мутации у дрозофилы. Из-за вполне понятной во время второй мировой войны секретности, результаты работы с этим отравляющим газом не были опубликованы до 1946 года. В том же 1946 году И.А. Рапопорт в СССР
продемонстрировал мутагенную
4-17

Глава 4. Изменчивость наследственного материала
активность формальдегида. С тех пор в |
В конце 1980х годов американские |
||||||||
арсенал мутагенных факторов вошли |
генетики А. Спрадлинг и Дж. Рубин |
||||||||
разнообразные химические соединения: |
предложили |
метод |
мутагенеза, |
||||||
аналоги оснований, включающиеся |
заключающийся в активировании |
||||||||
непосредственно |
â |
ДНК, такие |
перемещений мобильного P-элемента |
||||||
соединения как азотистая кислота или |
(см. раздел 6.6.), в результате чего он |
||||||||
гидроксиламин, |
|
|
соединения, |
может встраиваться в любой ген |
|||||
алкилирующие |
|
|
ÄÍÊ |
дрозофилы. |
Встройка |
(инсерция) |
|||
(этилметансульфонат, |
метилметан- |
мобильного |
элемента |
приводит к |
|||||
сульфонат |
è |
äð.), |
|
соединения, |
мутации данного гена. Таким образом по |
||||
интеркалирующие между основаниями |
морфологическим критериям можно |
||||||||
ДНК (акридины и их производные). Все |
отобрать мутантную линию дрозофил, в |
||||||||
ýòè |
вещества |
стали |
называть |
которой есть инсерция ДНК известного |
|||||
супермутагенами, из-за их высокой |
состава, что позволяет выделить ДНК |
||||||||
эффективности в индукции мутаций. Так |
мутировавшего гена (см. раздел ...). В |
||||||||
в работе И.А. Рапопорта 1946 года при |
результате исследователи дрозофилы |
||||||||
действии сублетальной дозы водного |
получили возможность выделять и |
||||||||
раствора |
формалина |
на личинок |
клонировать |
ÄÍÊ |
|
любого |
|||
дрозофилы было получено 47 летальных |
интересующего их гена. Это открытие |
||||||||
мутаций на 794 Х-хромосомы, изученных |
сделало революцию в молекулярной |
||||||||
по методу ClB (частота 5,9%), в контроле |
биологии. |
|
|
|
|||||
была найдена лишь одна летальная |
Изучение мутагенного действия |
||||||||
мутация на 833 хромосомы (частота |
ионизирующих излучений показало, что |
||||||||
0,12%). В работах Ауэрбах и Робсона |
у всех исследованных организмов они |
||||||||
частота мутаций достигала 24% (в |
вызывают многочисленные генные |
||||||||
контроле - 0,2%). (см. также Лобашев, |
мутации и перестройки хромосом и что |
||||||||
1967, ñ. 382-). |
|
|
|
частота индуцированных |
мутаций |
||||
В 1958 году С.И. Алиханяном и Т.С. |
зависит в основном от дозы радиации |
||||||||
Ильиной был установлен факт индукции |
(ðèñ. 4.8. a è á). |
|
|
||||||
мутаций у актиномицетов действием |
При этом не имеет большого |
||||||||
фагов. |
После |
этого |
появились |
значения, в один ли прием дана та или |
|||||
многочисленные |
публикации из |
иная доза или она разбита на дробные |
|||||||
многочисленных лабораторий, в которых |
порции, разделенные во времени - |
||||||||
установлено, что в результате вирусной |
мутагенный |
эффект |
â |
целом |
|||||
инфекции живых организмов (или клеток |
сооответствует общей дозе облучения. |
||||||||
в культуре) индуцируются хромосомные |
При облучении не существует нижнего |
||||||||
èëè |
хроматидные |
перестройки |
порога мутагенного действия (детали см. |
||||||
(транслокации, делеции, фрагментации |
Гершензон, 1983, стр. 226-240). |
||||||||
хромосом или их пульверизация), реже |
Первые работы по применению |
||||||||
анеуплоидия и полиплоидия. |
рентгеновских лучей в селекции были |
||||||||
4-18

Глава 4. Изменчивость наследственного материала
Табл. 4.7. Результаты селекции с применением мутагенных факторов у микроорганизмов-продуцентов некоторых антибиотиков (Из: Гершензон, 1991, стр. 94).
|
|
Активность (усл. ед.) |
||
Антибиотик |
Мутагены* |
|
|
|
Исходная |
Полученная |
|||
|
|
|||
|
|
|
|
|
Пенициллин |
Ð, ÓÔ, ÀÈ, ÝÈ |
220 |
5200 |
|
|
|
|
|
|
Стрептомицин |
ÀÈ, ÝÈ |
250 |
4200 |
|
|
|
|
|
|
Хлортетрациклин |
Ð, ÓÔ, ÝÈ |
600 |
2200 |
|
|
|
|
|
|
Эритромицин |
ÓÔ, ÝÈ |
500 |
2000 |
|
|
|
|
|
|
Альбомицин |
Ð |
2000 |
12000 |
|
|
|
|
|
|
Олеандомицин |
ÝÈ |
150 |
1500 |
|
|
|
|
|
|
* АИ - азотистый иприт, Р - рентгеновские лучи, УФ - ультрафиолетовые лучи, ЭИ - этиленамин.
|
Рис. 4.8.б Зависимость частоты |
|||
|
возникновениярецессивныхсцепленных |
|||
Рис. 4.8.а Прямолинейная зависимость |
с полом мутаций у дрозофилы (по оси |
|||
ординат в %%) от дозы рентгеновского |
||||
частоты видимых мутаций у |
||||
облучения (по оси абсцисс, Р). 1- |
||||
нейроспоры (по оси ординат Ч10-2) îò |
||||
теоретически |
ожидаемые, |
2- |
||
дозы рентгеновского облучения (по оси |
||||
абсциссЧ10000Р)(поданнымДемерека, |
экспериментальныеданные.(Поданным |
|
Тимофеева-Ресовского, см. Гершензон, |
||
см. Гершензон, 1983, стр. 228) |
||
1983, ñòð. 227). |
||
|
4-19

Глава 4. Изменчивость наследственного материала
проведены А.А. Сапегиным и Л.Н. Делоне в конце 1920-х - начале 1930-х годов, т.е. сразу же после открытия возможности искусственной индукции мутаций. Затем такие работы развернулись и в других странах, и в настоящее время эффективность экспериментального мутагенеза общепризнана.
Наиболее заметны успехи в селекции бактерий и грибов. Здесь быстрота смены поколений и огромное число особей в каждой культуре очень ускоряет темп селекции (Табл. 4.7).
В ряде случаев удалось повысить активность продуцентов в 10-20 раз, что позволило значительно увеличить производство соответствующих антибиотиков и резко снизило их стоимость, причем это было достигнуто в очень короткие сроки.
Похожие результаты получены при использовании мутагенов в селекции микроорганизмов, продуцирующих другие биологически активные вещества. Так, активность лучистого грибапродуцента витамина В12 - повысилась в 6 раз, а активность бактерии-продуцента аминокислоты лизина - даже в 300-400 раз (Из: Гершензон, 1991, стр. 93-94).
Литература к разделам 4.1. - 4.2.
Алиханян С.И., Ильина Т.С. Мутагенное действие актинофагов. Докл. Акад. Наук СССР, 120, 423-428, 1958.
Бородин П.М. Этюды о мутациях. Москва, Знание, I-III, 1983.
Вавилов Н.И. Закон гомологических рядов в наследственной измнчивости. (В кн. Классики советской генетики,
Ленинград, Наука, 9-50, 1968.) Гершензон С.М. Основы современной
генетики. Киев, Наукова Думка, 1- 558, 1983.
Гершензон С.М. Мутации. Киев, Наукова думка, I-III, 1991.
Инге-Вечтомов С.Г. Генетика с основами селекции. Москва, Высшая школа, 1- 592, 1989.
Лобашев М.Е. Генетика. Ленинград, Издво ЛГУ, 1-751, 1967.
Надсон Г.А., Филиппов Г.С. О влиянии рентгеновских лучей на половой процесс и образование мутантов у низших грибов (Mucoraceae). (В кн. Классики советской генетики, Ленинград, Наука, 120-124, 1968).
Gershenson S.M., Alexandrov Yu.N. Molecular mechanisms of mutagenecity of DNA and other natural and synthetic polynucleotides.PolymedPrinting,Kiev, 1-263, 1997.
Kilbey B.J. Charlotte Auerbach. Genetics 141, 1-5, 1995.
Lewin B. Genes V. Oxford, New York, Tokyo, Oxford University Press. p. 75, 1994.
Muller H.J. Artificial transmutation of the gene. Science 66, 84-87, 1927a.
Muller H.J. The problem of genic modification. Zeitschr. ind. Abst. Vererb. Sup.-Bd. 1, 234-260, 1927b.
Suzuki D.T., Kaufman T.C., Falk D and U.B.C. Drosophila Research Group. Conditionally expressed mutations in
Drosophila melanogaster. In: The genetics and biology of Drosophila, vol. 1a (M. Ashburner, E. Novitski, eds.)
London, New York, San Francisko, Academic Press, 208-263, 1976.
4-20