

Глава 7
ОСОБЕННОСТИ ОРИЕНТИРОВОЧНОЙ РЕАКЦИИ
КАК ДИАГНОСТИЧЕСКИЙ ПОКАЗАТЕЛЬ УРОВНЯ
СФОРМИРОВАННОСТИ НЕЙРОФИЗИОЛОГИЧЕСКИХ
МЕХАНИЗМОВ ВНИМАНИЯ И ПАМЯТИ У ДЕТЕЙ С НАРУШЕНИЯМИ ПОЗНАВАТЕЛЬНОЙ ДЕЯТЕЛЬНОСТИ
Проблемы развития, обучения и воспитания ребенка на разных этапах онтогенеза не могут быть успешно решены без знания специфики интегративной деятельности мозга ребенка и степени сформированности нейрофизиологических процессов, обеспечивающих реализацию высших психических функций.
Наблюдаемая в последние годы тенденция к увеличению количества детей, испытывающих трудности в обучении, является весьма актуальной проблемой. Преодолению этих трудностей должны способствовать научно обоснованные, адекватные коррекционные мероприятия, базирующиеся на эффективной, комплексной диагностике, направленной на выявление индивидуальных психофизиологических особенностей ребенка.
Современные клинико-физиологические и психолого-педагогические исследования популяции младших школьников показывают, что причиной трудностей обучения более чем у 50% неуспевающих учеников общеобразовательной школы является задержка психического развития (ЗПР); плохое усвоение знаний и представлений отмечается и у детей с общим недоразвитием речи (ОНР).
Важными в аспекте проблемы обеспечения дифференцированного подхода при обучении и воспитании детей с отклонениями в развитии представляются также исследования по изучению структуры дефекта в рамках одной формы аномалии.
В соответствии с клиническими и психофизиологическими данными особенности познавательной деятельности, присущие детям с ЗПР, в значительной мере определяются уровнем сформированности регуляторных и познавательных про-
238
цессов. Это позволяет выделить из полиморфного контингента детей с ЗПР две основные группы. Одну из них (более 30% детей с ЗПР) составляют дети, у которых ведущим фактором в структуре отклонений в психическом развитии является недостаточная сформированность регуляторных процессов, проявляющаяся в дефиците внимания и особенностях эмоционально-волевой сферы, что в значительной мере компенсируется в процессе возрастного развития; у детей с такими особенностями недостатки в развитии памяти, вербального и невербального мышления могут иметь вторичный характер. Для другой, значительной части детей с ЗПР (около 40%) ведущим фактором в структуре дефекта является недостаточность познавательных процессов (памяти, восприятия, мышления), которая может иметь стойкий характер (Диагностика и коррекция..., 1988; М.С. Певзнер, 1972, 1982; Л.И. Переслени, 1984; Л.И. Переслени, Е.М. Масткжова, 1989). Сходные результаты были получены и при исследовании особенностей познавательных процессов у детей с ОНР (Л.И. Переслени, Т.А. Фотекова, 1993).
Следует отметить, что по классификации, используемой западными исследователями, выделяемой нами первой группе детей в наибольшей степени соответствуют дети с нарушениями, обусловленными дефицитом внимания (Attention Deficit Disorder), среди которых, в свою очередь, выделяются две подгруппы с или без гиперактивности. Вторая группа детей может быть соотнесена с обширной, более гетерогенной по структуре нарушений познавательной деятельности категорией детей с трудностями обучения (Learning disabilities).
В последние годы в ряде исследований подчеркивается сложность дифференциальной диагностики указанных форм аномального развития (ЗПР и ОНР), поскольку их отдельные клинико-психофизиологические проявления сходны, а иногда совпадают (Л.И. Переслени, Т.А. Фотекова, 1993; С.Г. Шевченко, 1994; Т.Н. Волковская, 1999). Это позволяет выделять случаи сложного дефекта, когда ЗПР сочетается с ОНР. И в западной литературе отмечается, что среди школьников, испытывающих трудности в обучении, свыше 90% имеют речевые и языковые нарушения, при этом в 20-35% случаев у таких детей обнаруживается и дефицит внимания в сочетании с гиперактивностью; и наоборот, 50% детей с дефицитом внимания имеют также трудности в обучении и другие академические проблемы (L.B. Silver, 1990; A. Talay-Ongan, 1998).
239
Несмотря на патогенетическую неоднородность детей с ЗПР и ОНР, представляется целесообразным и важным выделять среди них две основные подгруппы, для одной из которых характерна недостаточность преимущественно регуляторных функций (внимание, эмоционально-волевая сфера), а для другой - познавательных процессов (восприятие, память, мышление). Характер соотношения между уровнем развития регуляторных и познавательных процессов важно учитывать при диагностике не только интеллектуальных, но и сенсорных дефектов (нарушения зрительного, слухового восприятия), поскольку и они часто сочетаются с недостаточностью актива-ционных, регуляторных и мнестических процессов (Э.С. Опо-линский, Л.А. Рожкова, 1993; Л.П. Григорьева, 2000).
Обоснованность указанного подхода для оценки состояния познавательных процессов опирается на представления о системной организации деятельности мозга (А.Р. Лурия, 1973) и данные значительного числа психофизиологических и нейрофизиологических исследований (Д.А. Фарбер, 1978; Л.И. Пе-реслени, 1984; Л.И. Переслени, Е.М. Мастюкова, 1989; Е.Д. Хомская, 1987, 1999; В.И. Лубовский, 1989; Л.П. Григорьева и др., 1997).
Научно обоснованная разработка и выбор наиболее эффективных коррекционных мероприятий для компенсации отставаний в развитии детей с трудностями обучения различной этиологии невозможны без понимания глубинных причин выявляемых нарушений познавательной деятельности ребенка.
Многочисленные нейроанатомические, нейрохимические, нейрофизиологические данные свидетельствуют о том, что нарушения, отклонения в развитии необходимых для усвоения знаний и навыков психических функций у детей с трудностями обучения во многих случаях имеют неврологическую основу. При этом специфика, степень выраженности поведенческих проявлений нарушения познавательной деятельности определяется качественным своеобразием, сложностью лежащего в их основе неврологического субстрата (М.С. Певзнер, 1972; Е.М. Мастюкова, 1990; Л.И. Переслени и др., 1996; L.B. Silver, 1990; K.K.S. Voeller, 1991; С.А. Riccio et al., 1993). Так, нарушения внимания могут быть связаны с дисфункцией передних (лобных) или задних (теменных) отделов коры, базальных ганглиев, стволовых структур вследствие специфики (незрелость или аномалия) их морфофункциональной организации, в ряде случаев наиболее выраженной в правом полушарии. Кроме
240
того, при дефиците внимания и нарушениях памяти отмечается дисбаланс в образовании катехоламинов - одной из основных групп нейромедиаторов (Е.А. Громова, 1980; K.K.S. Voeller, 1991; G. Hynd et al., 1993; С.А. Riccio et al., 1993; A. Talay-Ongan, 1998). Различные сочетания вовлечения в патологический процесс тех или иных структур мозга, в частности, передних (лобных) или задних (теменных) отделов коры коррелируют с различными поведенческими проявлениями: например, дефицит внимания с или без гиперактивности (P. Goodyear, G.W. Hynd, 1992; С.А. Riccio et al., 1993; V.R. Brever et al., 1997). Вместе с тем, в настоящее время нет единых представлений о нейробиологической основе указанных нарушений познавательной деятельности. По одним данным, в основе нарушений внимания, сочетающимися с гиперактивностью, лежит ослабление стволовых активирующих влияний (D. Cantwell, 1977; J.H. Satterfield, M.E. Dawson, 1971; S.S. Zentall, 1993), по другим - дефицит кортикофугальных тормозных влияний (R.H. Rosentall, T.W. Allen, 1978; R.A. Dykman, P.T. Ackerman, 1993; A. Talay-Ongan, 1998).
Нейрофизиологические исследования дают возможность объяснить динамическое взаимодействие между нейрохимическими и нейроанатомическими компонентами, образующими функциональную систему. Изучение особенностей взаимодействия и взаимовлияния различных структур мозга при осуществлении того или иного вида деятельности может способствовать пониманию нейробиологической основы трудностей обучения различной этиологии. Такие исследования представляются важной, высоко значимой составляющей при реализации системного подхода к изучению нарушений развития, необходимость применения которого подчеркивалась еще Л.С. Выготским (1983).
Большое значение в аспекте вышеизложенного имеет изучение нейрофизиологических механизмов процессов активации и регуляции и в особенности - степени ссрормированности нисходящих регулирующих влияний со стороны лобных отделов коры головного мозга на разных этапах онтогенеза, поскольку в соответствии с клиническими и нейрофизиологическими данными для значительной части детей с ЗПР (с психофизическим и органическим инфантилизмом) характерно недоразвитие лобных структур и их связей (М.С. Певзнер, 1972, 1982; И.Ф. Марковская, 1993; Л.И. Переслени, Е.М. Мастюкова, 1989; М.Н. Фишман, 1989). Последнее является причиной
241
незрелости произвольной регуляции деятельности, проявляющейся в особенностях произвольного внимания и эмоционально-волевой сферы. В связи с длительным периодом созревания этих отделов коры в онтогенезе (М.С. Певзнер, 1982; Е.П. Кононова, 1965; Физиология подростков, 1988) высказывались предположения о том, что по мере морфофункцио-нального развития мозга эти отклонения могут компенсироваться в старшем школьном возрасте.
Несформированность процессов активации мозговых структур и механизмов корковой регуляции, по мнению ряда авторов, может служить нейробиологической основой нарушения речевого развития (M.S. Gazzaniga, 1974; М. Hiscock, М. Kinsbourne, 1980, 1987; М. Kinsbourne, 1981; J.R. Kershner, R.W. Stringer, 1991). Так, в соответствии с гипотезой М. Kinsbourne, речевой дизонтогенез может быть обусловлен задержкой созревания нервного субстрата, в частности, структур левого полушария, дифференцированная специфическая активация которого в норме обеспечивает адекватную реализацию речевой функции. M.S. Gazzaniga выдвигает предположение о том, что нарушения речевого развития могут определяться незрелостью системы контроля над психическими процессами, нейрофизиологической основой которого являются тормозные механизмы.
Одним из наиболее важных аспектов проблемы изучения нарушений познавательной деятельности представляется исследование мозговых механизмов таких базисных процессов как внимание и память, которые в значительной степени определяют закономерности протекания любой психической функции. Внимание и мнестические процессы, в свою очередь, непосредственно связаны с уровнем сформированности корковой ритмики, степенью развития процессов регуляции и следообразования. При этом особенности стволовых активирующих и регулирующих корковых влияний, являющихся нейрофизиологической основой непроизвольного и произвольного внимания, в значительной мере обусловливают и специфику процессов запечатления и воспроизведения информации, что находит отражение в особенностях возникновения и угашения ориентировочной реакции (ОР). Интенсивность и распространенность активирующих влияний в системе корково-подкорковых связей модулируются механизмами регулируемой корковой активации, что способствует селективности восприятия информации и в значительной степени
242
![]()
определяет эффективность процессов следообразования и актуализации следов. Реализация последних осуществляется через циклические процессы, самым тесным образом связанные с ритмической активностью мозга. Отсюда вытекает важность изучения степени сформированности корковой ритмики различных частотных диапазонов и в особенности ее реорганизации в процессе поступления сенсорных стимулов различной информационной значимости.
Проводимые нами в последние годы нейропсихофизиоло-гические исследования направлены на изучение особенностей функционального состояния ЦНС и нейрофизиологических механизмов восприятия информации у детей с ЗПР и ОНР. Они показали информативность изучения указанных составляющих интегративной деятельности мозга для понимания структуры дефекта и подтвердили обоснованность представленного выше подхода к дифференциальной диагностике. Было показано, что соотношение уровня развития регуляторных и познавательных процессов, выявляемое в ходе психологического обследования ребенка и, в частности, при анализе прогностической деятельности, в значительной степени коррелирует с характеристиками развития электроактивности коры мозга, состоянием стволовых регуляторных структур и главное, с особенностями ОР (Л.И. Переслени, Л.А. Рожкова, 1990, 1991, 1993, 1996; Л.И. Переслени, Л.А. Рожкова, Н.А. Рябчикова, 1990; Л.И. Переслени, Л.А. Рожкова, Т.А. Фотекова, 1994; Л.А. Рожкова, 1997, 2000; L.I. Peresleni, L.A. Rozhkova, N.A. Ryabchikova, 1991). Это позволяет использовать характеристики компонентов ОР как дополнительный, объективный критерий оценки уровня сформированности активирующих и регулирующих влияний, а также процессов следообразования. Этот метод приобретает особую значимость в диагностике нарушений внимания, что является достаточно сложной задачей (K.K.S. Voeller, 1991; К. McBarnett et al., 1993; Н.М. Borchgrevinc, 1997; A. Talay-Ongan, 1998). Так, в ряде случаев особенности поведенческих реакций ребенка в процессе психологического обследования, связываемые с нарушениями внимания, могут быть обусловлены не столько функциональной недостаточностью систем активации и регуляции, сколько особенностями отношения ребенка к выполняемому заданию (недостаточный интерес, сниженная мотивация и т.п.), спецификой контакта с психологом, педагогом, исследователем. С другой стороны, нарушения внимания могут существенно вли-
243
ять на эффективность мнестических процессов, в значительной степени детерминируя результаты тестов на память. Кроме того, появляется все больше доказательств цитоархитекто-нической дифференциации внутри фронтальной коры, что обусловливает ее важную роль в реализации не только регу-ляторных, но и мнестических процессов (поиск, воспроизведение информации) (Функции лобных долей мозга, 1982; М.А. Wheeler et al., 1995).
В связи с этим в дополнение к общепринятому клиническому методу регистрации "фоновой" ЭЭГ, позволяющему оценить функциональное состояние мозговых структур в "покое" и при функциональных нагрузках, предлагается ввести в практику клинического обследования детей с трудностями обучения различного генеза регистрацию и анализ особенностей возникновения и угашения ориентировочной реакции (ОР) на сенсорные стимулы различной сигнальной значимости.
Феномен ориентировочного рефлекса был впервые описан выдающимся русским физиологом И.П. Павловым (1952) как реакция организма на внезапное изменение окружающей среды, имеющая большое биологическое значение и названа ориентировочно-исследовательской или реакцией "Что такое?".
Многочисленные нейропсихофизиологические исследования показывают, что ОР есть результат активности весьма сложной функциональной системы, видоизменяющей деятельность многих других функциональных систем организма, о чем свидетельствует многофакторное проявление ориентировочной реакции: электроэнцефалографические компоненты ОР (реакция активации, неспецифический ответ (НО), позднее позитивное колебание РЗ) регистрируются наряду со многими вегетативными сдвигами (изменение ритма сердца, дыхания, плетизмографическая реакция сосудов, кожно-галь-ванический рефлекс (КГР) и др.) (П.К. Анохин, 1975; Л. А. Новикова и др., 1959; Е.Н. Соколов, 1964, 1969; Л.П. Латаш, 1968; Р. Наатанен, 1998; И.М. Фейгенберг, 1998; N. Lovless, 1983: R. Naatanen, 1986). Среди паттерна эффектов ОР Р. Наатанен (1998) выделяет два основных компонента: энергетический, или активационный, необходимый для активации сенсомотор-ных функций, и информационный, связанный с рефлекторным включением внимания.
Одним из наиболее мощных возбудителей ОР является "новизна" стимула, определяющаяся степенью несовпадения
244
![]()
поступающего сигнала с "нервной моделью стимула" (НМС), эталоном, сформированным в нервной системе. Поэтому наиболее отчетливо она проявляется при первом, неожиданном предъявлении афферентного сигнала, а затем постепенно угасает (Е.Н. Соколов, 1969) и рассматривается как коррелят непроизвольного внимания (Р. Наатанен, 1998). Наряду с этим ОР вызывается когнитивным рассогласованием (например, при внезапном повторении предыдущего сигнала в ряду чередующихся различных стимулов), а также усиливается при повышении значимости стимула, т.е. в условиях произвольного внимания (Р. Наатанен, 1998; R. Naatanen, 1986).
Развитие и угашение ОР, составляющие процесс привыкания, обеспечиваются взаимодействием иерархии центральных структур мозга, в число которых наряду с сенсорными системами входят ретикулярные образования, а также ассоциативные системы мозга.
Одной из особенностей ОР является эффект ее растор-маживания, проявляющийся в восстановлении реакции на раздражитель, на который ОР была угашена, после применения нового агента. Этот эффект объясняется деблокированием путей, ведущих к ядрам ретикулярной формации, являющимся центром эфферентной интеграции ОР (Е.Н. Соколов, 1964, 1969).
При одновременной регистрации разных компонентов ОР было обнаружено, что их угашение при повторных предъявлениях индифферентного стимула происходит не одномоментно и отличается различной устойчивостью. По скорости угашения регистрировавшиеся компоненты могут быть расположены (в порядке убывания скорости, т.е. замедления угашения) следующим образом: изменения частоты сердечного ритма-изменения дыхания-КГР—активация ЭЭГ—неспецифический ответ (НО). При угашении ОР на значимый стимул этот порядок меняется таким образом, что наиболее медленно угашаемым становится КГР и повышается угашаемость ЭЭГ-компонентов реакции: НО - активация ЭЭГ-КГР (Л.П. Латаш, 1968).
Реакция активации (или arousal - пробуждение) рассматривается как корковый компонент ОР, проявляющийся в десин-хронизации, угнетении ритмической активности мозга. Она является отражением активирующих влияний ретикулярной формации среднего мозга и зависит от степени зрелости и сохранности корковых нейронов. Поэтому характеристики ре-
245
акции активации изменяются в онтогенезе, а также коррелируют с особенностями корково-подкорковых взаимодействий при отклонениях в развитии той или иной этиологии.
Выраженность реакции активации (как и других компонентов ОР) зависит от информационной или биологической значимости стимулов.
Привлечение с помощью инструкции внимания к ранее индифферентным раздражителям, вовлекающее в процесс восприятия переднеассоциативные отделы коры, изменяет не только интенсивность реакции десинхронизации, но и специфику соотношения ее выраженности в различных областях коры. Анализ этих характеристик дает возможность оценить степень сформированности или сохранности кортикофугаль-ных регулирующих влияний, а также других системных процессов, обеспечивающих осуществление функции внимания (Н.В. Дубровинская, 1985; Л.И. Переслени, Л.А. Рожкова, 1990, 1991, 1993, 1996; Л.А. Рожкова, 1997; Л.П. Григорьева и др., 1997; К.Н. Rosenthal, N.W. Allen, 1978).
В последние годы в связи с усовершенствованием методов обработки и анализа биоэлектрических сигналов возрос интерес к изучению различных процессов, развивающихся в период протекания вызванной стимулом десинхронизации ЭА, что позволяет получить новые сведения о механизмах и функциональной значимости этого ЭЭГ-феномена (В.А. Пономарев и др., 1999; Ю.Д. Кропотов и др., 1999; G. Pfurtsheller, W. Klimesch, 1989).
Вегетативный компонент ОР-КГР характеризует уровень эмоционального напряжения, связанный с состоянием эмоци-огенных структур мозга как в условиях спокойного бодрствования, так и при осуществлении любой целенаправленной деятельности (Л.П. Латаш, 1968). Показана прямая связь между эффективностью мнестической деятельности и величиной, а также реактивностью КГР (Э.А. Громова, 1980; Н.В. Макаренко и др., 1988). Однако существует оптимальный для когнитивной деятельности уровень эмоционального напряжения, характеризуемый умеренной активацией ЦНС.
Динамика изменения выраженности электрокорковых и вегетативных компонентов ОР при повторении стимулов характеризует процесс привыкания (или "негативного научения") и является косвенным отражением процесса формирования НМС, т.е. следообразования, базирующегося на ассоциативном механизме мозга (Е.Н. Соколов, 1969; Б.И. Котляр,
246
1989; R. Naatanen, 1986). Это позволяет использовать характеристики угашения ОР как дополнительный критерий оценки состояния некоторых базисных нейрофизиологических механизмов мнестической функции.
В современных исследованиях в качестве ЭЭГ-корреля-та протекания ряда когнитивных процессов часто рассматривается позднее позитивное колебание, возникающее в ряду компонентов связанного со стимулом потенциала в тех случаях, если стимул является новым, неожиданным или обладает какой-либо информационной значимостью для испытуемого. Этот компонент достигает амплитудного максимума через ЗОО^ЮО мс после предъявления сигнала и обозначается как РЗОО или РЗ (Э.М. Рутман, 1979; Э.С. Ополин-ский, Л.А. Рожкова, 1990; Р. Наатанен, 1998; N. Lovless, 1983; С.С. Wood et al., 1984).
Таким образом, исследование особенностей возникновения и угашения ОР занимает весьма важное место в ряду физиологических проблем и может способствовать пониманию нейрофизиологических механизмов, лежащих в основе инте-гративной деятельности мозга и обеспечивающих осуществление высших психических функций как в норме, так и при отклонениях в развитии различной этиологии. Использование условий непроизвольного и произвольного внимания дает возможность оценить особенности состояния, участия в реализации ОР не только восходящих стволовых активирующих влияний, но и системы корковой регулируемой активации. Именно анализ изменений характеристик ОР при повышении значимости стимулов позволяет выявить наиболее существенные отличия детей с разными формами аномального развития от нормы по уровню сформированности этих системных процессов и представляет наибольшую дифференциально-диагностическую ценность.
7.1. Метод исследования
Обследовались дети, которые по данным ПМПК характеризовались ЗПР или ОНР.
При ЗПР исследовались дети трех возрастных групп: младшего (6,5-9 лет), среднего (9,5-12 лет) и старшего (12,5-15 лет) школьного возраста (всего 80 чел.).
Группу с речевой патологией составили дети с ОНР II-III уровня только младшего школьного возраста (68 чел.).
247
Среди детей с ЗПР и ОНР были выделены по две подгруппы, одна из которых в соответствии с педагогическими характеристиками и данными психологического обследования (решение прогностической задачи по методике "Угадайка" -Л.И. Переслени, В.Л. Подобед, 1982) характеризовалась преимущественно дефицитом внимания (ДВ), а другая - дефицитом памяти (ДП).
Контрольную группу (40 чел.) составили учащиеся общеобразовательных школ младшего, среднего и старшего школьного возраста, которые имели хорошую успеваемость и по показателям прогностической деятельности характеризовались сформированностью процессов внимания и памяти.
Электрофизиологическое обследование, направленное на оценку функционального состояния ЦНС и особенностей ОР при восприятии сенсорных стимулов различной сигнальной значимости, проводилось в следующих ситуациях:
- в покое (спокойное бодрствование);
- при предъявлении 15 световых вспышек с нерегулярными интервалами (межстимульный интервал 2-3,5 сек.) в условиях: а) непроизвольного внимания (НПВ) (угашение ОР); б) произвольного внимания (ПВ), когда испытуемого просят сосчитать количество стимулов в серии;
- при функциональных нагрузках (ритмическая фотостимуляция и гипервентиляция). Общая длительность электрофизиологического исследования составляет около 30 мин.
Испытуемые при обследовании находятся в экранированной, звукозаглушенной камере, сидя в кресле в темноте с закрытыми глазами.
ЭЭГ регистрируется стандартными электродами, расположенными в соответствии с Международной системой 10/20, от симметричных точек ряда областей правого (d) и левого (s) полушарий головного мозга: затылочных (О), теменных (Р), центральных (С), лобных (F), передне- (Та) и задне-височных (Тр), а также от центральной и лобной сагиттальных зон (Cz и Fz). Референтными служат ушные электроды. Параллельно с ЭЭГ регистрируется КГР по Тарханову - с левой руки. Усиление сигналов осуществляется с помощью многоканального энцефалографа фирмы "Nihon Kohden" (Япония); полоса пропускания от 0,1 до 30 Гц. Предъявление сенсорных стимулов может осуществляться как в режиме ручной стимуляции (с панели электро-
248
энцефалографа), так и автоматически с помощью цифрового компьютера, который управляет (в соответствии с созданными схемами стимуляции) световым стимулятором (лампа-вспышка), оцифровывает аналоговые сигналы (частота опроса 125 Гц) и визуализирует процесс регистрации в режиме on-line, а также накапливает данные для дальнейшей обработки. Эпоха анализа составляет 30 сек. при регистрации фоновой ЭЭГ и 2000 мс при регистрации ОР.
Для регистрации и обработки ЭЭГ нами использовался аппаратно-программный комплекс "Нейрокартограф", разработанный научно-медицинской фирмой МБН (Москва).
Для каждого обследуемого целесообразно анализировать следующие электрофизиологические характеристики:
- спектральная плотность мощности- (СПМ в мкв/Гц) основных ритмов фоновой ЭЭГ 14 областей коры в следующих частотных диапазонах: тета 1 (3,7-5,5 Гц), тета 2 (5,7-7,4), альфа 1 (7,6-8,9), альфа 2 (9,1-12,9), бета 1 (13,1-19,9) и бета 2 (20-30 Гц). По данным спектрального анализа осуществляется картирование основных ритмов ЭЭГ;
- наличие и характер ЭЭГ-знаков, указывающих на дисфункцию или повреждение различных структур мозга;
- выраженность КГР при стимуляции (среднее количество реакций по отношению к общему числу предъявленных стимулов в процентах), а также наличие КГР в фоновой записи;
— амплитудные (А) и временные (пиковая латентность — ПЛ) характеристики компонента РЗ в зрительном ВП, полученном при усреднении вызванной активности на первые три стимула. Для оценки этих показателей использовалась специально разработанная программа обработки ЭА мозга, которая вычисляет интегральную амплитуду (А инт.) колебаний (измеренных относительно нулевой линии, определяемой по 304 мс отрезка престимульной активности) в выделенном временном интервале ("окне") Т; интервалы (соответствующие колебанию РЗ, а также реакции десинхрони-зации (РД)) выделяются при визуальном анализе на экране дисплея индивидуальных, связанных со стимулом усредненных реализаций ЭА (отдельно для совокупности каудаль-ных О, Р, Тр и передних С, F, Та областей). В связи с индивидуальной вариабельностью длительности Т, мы оценивали средние значения А: А ср. = А инт/Т;
- длительность реакции десинхронизации (ДРД) в мс в затылочной и центральной областях коры левого полушария,
249
оцениваемая визуально по среднему арифметическому значению на первые три стимула в ситуациях 2а и 26.
В целях оптимизации обследования визуальный анализ длительности реакции активации осуществляется лишь для проекционной и ассоциативной областей коры левого полушария в связи с имеющимися данными о специфике вовлечения его структур в процесс решения непространственных задач, а также учитывая важную роль левого полушария в осуществлении речевых функций.
Использовался также нетрадиционный метод анализа, позволяющий оценить А ср. (а также полярность) колебаний в период Т, связанный с десинхронизацией корковой ритмики в ответ на предъявление сенсорного стимула. Этот показатель вычислялся аналогично А ср. для компонента РЗ и обозначен как АРД ("амплитуда" реакции десинхронизации). Он отражает величину и направленность (минус - негативация, плюс -позитивация) сдвигов ЭА различных областей коры в период, связанный с РД.
Результаты нейрофизиологического исследования соотносили с поведенческими характеристиками различных психических функций (внимание, память, мышление), выявляемыми в ходе психологического обследования.
Для оценки статистической достоверности полученных результатов использовался непараметрический критерий Т Вил-коксона (при анализе ДРД и СПМ), а также методы дисперсионного анализа (ANOVA/MANOVA, пакет программ STATISTICA). При этом оценивалось влияние на амплитудно-временные характеристики электрокорковых компонентов ОР следующих факторов: функциональное состояние (ФС: НПВ/ПВ), объект (норма/ОНР/ДВ//ОНР/ДП).
Совокупность исследуемых показателей (спектральные характеристики фоновой ЭЭГ, выраженность знаков дисфункции или повреждения различных структур мозга, характеристики компонента РЗ и реакции десинхронизации на индифферентные и релевантные зрительные стимулы) позволяют оценить степень сформированности корковой ритмики, активирующих и регулирующих влияний, в значительной степени определяющих специфику реализации высших психических функций, что может способствовать пониманию и прогнозу особенностей когнитивных нарушений при ЗПР и ОНР.
250
7.2. Характеристики компонентов
ориентировочной реакции у нормально развивающихся школьников
В данном разделе рассматриваются характеристики компонентов ОР на индифферентные и значимые сенсорные стимулы у детей в норме младшего школьного возраста (6,5-9,0 лет), а также анализируются некоторые их возрастные изменения в среднем (9,5-12,0 лет) и старшем (12,5-15,5 лет) школьном возрасте.
А. Условия непроизвольного внимания. Анализ амплитудно-временных и пространственных характеристик компонента РЗ в ВП, связанных с предъявлением первых трех вспышек в серии (т.е. при наибольшей выраженности ОР), у младших школьников показал следующее. Амплитудный максимум РЗ отмечается в теменных отделах (T-test, p < 0,01), при этом пиковая латентность (ПЛ) в теменных и затылочных отделах наименьшая (р < 0,05 в сравнении с Тр для левого полушария); в правом полушарии только в центральной области ПЛ РЗ существенно меньше, чем в Тр (р < 0,09). Следует отметить, что в лобных отделах в этом временном периоде отмечается негативация ЭА (рис. 7.1-II).
Межполушарная асимметрия РЗ выражена только в затылочной коре и проявляется в левополушарном преобладании его амплитуды (/; < 0,05).
Вслед за РЗ обычно развивается реакция активации, проявляющаяся в виде десинхронизации электроактивности мозга, которая наиболее отчетливо регистрируется при первых двух-трех предъявлениях сенсорного раздражителя (рис. 7.1). Анализ длительности реакции активации показал, что в условиях непроизвольного внимания она больше в затылочной, проекционной коре по сравнению с центральной, ассоциативной зоной (рис. 7.1; табл. 7.1). Такое соотношение длительности РД при поступлении индифферентных стимулов отмечалось нами на всех этапах возрастного развития (младший, средний, старший школьный возраст).
У младших школьников реакция десинхронизации характеризуется неустойчивостью: после предъявления первых трех стимулов она выявляется нерегулярно, что делает нецелесообразным анализ динамики ее угашения. В среднем и старшем школьном возрасте по мере морфофункциональ-ного созревания структур мозга и усиления активирующих
251
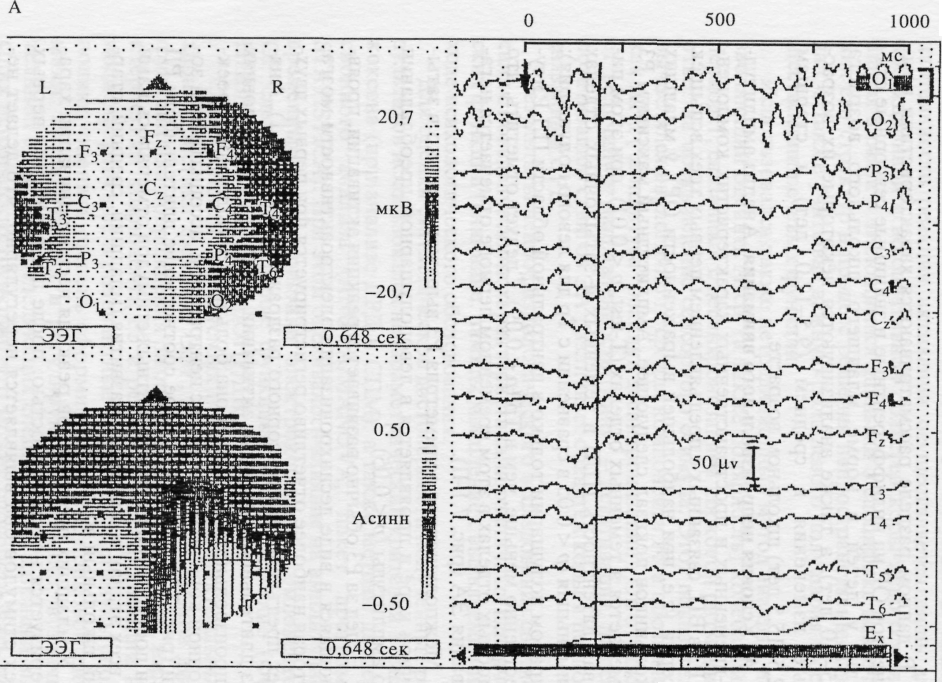
Рис. 7.1 (I, А). Подрисуночный текст на стр. 255

Рис. 7.1 (I, Б) (продолжение)



Рис. 7.1 (II, А, Б, В) (окончание)
влияний со стороны ретикулярной формации ствола усиливается и реакция активации, что находит отражение в увеличении ее длительности, повышении регулярности появления. Использование нетрадиционного метода анализа (помимо визуальной оценки ДРД) позволило оценить степень и полярность сдвигов биопотенциалов в период, связанный с наиболее выраженной десинхронизацией корковой ритмики в ответ на предъявление стимула, что дало возможность
254
Таблица 7.1
Особенности реакции десинхронизации электроактивности головного мозга у детей в норме
ДРД (мс)
|
Возраст |
Затылочная обл. (Os) |
Центральная обл. (Cs) | ||||||
|
непроизвольное внимание |
произвольное внимание |
непроизвольное внимание |
произвольное внимание | |||||
|
6,5-9,0 |
650 |
560 |
530 |
760 | ||||
|
9,5-12,0 |
650 |
670 |
480 |
750 | ||||
|
12,5-15,5 |
960 |
700 |
640 |
1040 | ||||
выявить регионарную специфичность этого показателя и его существенную связь с уровнем внимания.
Установлено, что у младших школьников в ситуации непроизвольного внимания в период развития РД в затылочной коре, особенно справа, отмечается выраженный сдвиг ЭА в сторону негативности (негативация) (р < 0,05). В других отделах, главным образом в переднеассоциативных (F, С, Та), напротив, наблюдается позитивация ЭА (рис. 7.1-II, В). Очевидно, можно говорить о локальной негативации ЭА в проекционных зонах коры зрительного анализатора в сочетании с генерализованной позитивацией биопотенциалов в других отделах мозга при неожиданном поступлении новой информации.
Рис. 7.1. Особенности электрокорковых компонентов ОР у детей в норме.
I: компонент РЗОО и реакция десинхронизации корковой ритмики у испытуемого Р.Ю., 7 лет в различных областях коры мозга при предъявлении первого светового стимула в условиях ненаправленного (А) и направленного (Б) внимания. Слева представлены топографические карты распределения по скальпу (верхняя карта) и межполушар-ной асимметрии (нижняя карта) амплитудных значений компонента РЗОО в момент времени, помеченный курсором (вертикальная линия). Стрелкой отмечен момент предъявления стимула. О - затылочные, Р - теменные, С - центральные, F - лобные, Т3,4 - передневисочные, Т5,6 - задневисочные отведения левого (L, нечетные цифры) и правого (R, четные цифры) полушарий, Cz и Fz - сагиттальные отведения; Exl - КГР.
II: среднегрупповые значения амплитуды (А) и латентности (Б) компонента РЗОО, величины сдвига ЭА в период РД (В) в различных зонах коры в условиях непроизвольного и произвольного внимания у детей в норме
255
Следует отметить, что различия величин потенциалов в период развития РЗ и РД достоверны во всех областях, кроме височных (р < 0,04). Однако, если в затылочно-теменных и центральных отделах эти различия характеризуют сдвиг от позитивности РЗ к большей негативности в период РД (или уменьшение позитивации), то в лобных отделах изменения носят противоположный характер - от негативации в период РЗ к позитивации при РД (р < 0,05).
КГР в состоянии покоя регистрировалась у 62% детей. При стимуляции она возникала на первые 5-6 стимулов, т.е. отмечалась в 30% случаев.
Б. Условия произвольного внимания. Степень выраженности электрокорковых компонентов ОР, особенности их межрегионального распределения зависят также от информационной значимости воспринимаемых раздражителей.
Привлечение внимания к стимулам (которое осуществлялось нами во второй серии, т.е. после угашения ОР, связанной с новизной стимула) находит отражение лишь в тенденции к увеличению амплитуды РЗ, наиболее выраженной в затылочной и передневисочной зонах правого полушария (рис. 7.1-П). Амплитудный максимум РЗ сохраняется в теменных зонах (р < 0,001 при сопоставлении с С, F, Та, Тр); наименьшая позитивность отмечается в передневисочных зонах.
Наиболее существенно повышение уровня внимания влияет на ПЛ РЗ: она уменьшается во всех областях (рис.7.1-Й, Б; р < 0,05 для симметричных затылочных и задневисочных зон коры, левой передневисочной и правой теменной). При этом в отличие от условий непроизвольного внимания межрегиональные различия значений ПЛ РЗ становятся менее выраженными, но минимальная ее величина выявляется в затылочных зонах обоих полушарий (р < 0,04 в сравнении с Та, Тр).
Межполушарная асимметрия РЗ также становится менее выраженной, чем в ситуации непроизвольного внимания, и достоверно не проявляется.
ДРД у детей всех возрастных групп при повышении значимости стимулов в затылочной области коры уменьшается (особенно у старших школьников). Напротив, ДРД в центральной корковой зоне на релевантные стимулы всегда и существенно больше ее значений на индифферентные сигналы, причем с возрастом эти различия усиливаются и составляют 43, 56 и 62, 5% в трех рассматриваемых возрастных группах соответственно (см. табл. 7.1).
256
Следует подчеркнуть, что привлечение внимания к стимулам изменяет и соотношение степени вовлечения в ОР проекционных и ассоциативных зон коры. Так, в отличие от условий непроизвольного (ненаправленного) внимания, когда ДРД в затылочной коре больше, чем в центральной, при активном анализе сигналов этот показатель в центральной области становится значительно больше, чем в затылочной. Такое соотношение выявлено нами на всех исследованных этапах возрастного развития, но в наибольшей степени оно выражено у старших школьников.
Угашение реакции активации проявляется в уменьшении ее длительности, которая после предъявления 3-4 стимулов относительно стабилизируется у детей среднего и старшего школьного возраста. Следует отметить, что декремент ДРД на релевантные сигналы в центральной области коры с возрастом увеличивается.
Привлечение внимания к стимулам вызывает выраженные изменения не только длительности реакции активации, но и других характеристик ЭА в период ее развития. Установлено изменение направленности сдвигов ЭА на противоположную в сравнении с ситуацией непроизвольного внимания, а именно негати-вация ЭА всех зон коры (фактор ФС: р < 0,05, кроме Та левого полушария и Р и Тр - правого) за исключением затылочных отделов, где отмечалось усиление негативации (рис. 7.1-П, В). При этом в левом полушарии максимум негативации отмечается в лобной коре (р < 0,05 в сравнении с центральной зоной), а в правом - в затылочной (р < 0,05 в сравнении с Р, С, Та, Тр). Отметим, что, как и в ситуации ненаправленного внимания, межполу-шарные различия данного показателя наиболее выражены в проекционной коре и характеризуются большей негативацией затылочной зоны правого полушария (р < 0,05).
Сравнение величины РЗ с характеристиками ЭА в период РД выявляет следующую закономерность: в ситуации непроизвольного внимания, т.е. при восприятии новых, неожиданных сигналов большей амплитуде РЗ соответствует меньшая негативация в период РД; привлечение внимания способствует как росту амплитуды РЗ, так и, в особенности, усилению негативности в период РД, что увеличивает градиент потенциалов между РЗ и РД (р < 0,001 для О, Р, С, Тр). Только в лобных отделах этот градиент сглаживается по сравнению с условиями непроизвольного внимания вследствие негативации ЭА в период развития и РЗ, и РД.
257
Выраженность КГР также обнаруживает зависимость от значимости воспринимаемой информации. Так, частота встречаемости КГР при предъявлении релевантных сигналов выше на 20% (т.е. скорость угашения меньше) по сравнению с таковой на иррелевантные стимулы. Этот показатель также увеличивается с возрастом.
Кроме того, по мере возрастного развития отмечается уменьшение частоты встречаемости КГР в состоянии спокойного бодрствования, что отражает формирование системы регуляции, обеспечивающей оптимальный уровень активации эмоциогенных структур мозга.
Следовательно, рассмотренные показатели обнаруживают выраженную корреляцию с процессами неспецифической активации и корковой регуляции при восприятии сенсорной информации. Поскольку представленный анализ изменений ЭА в период развития РД является нетрадиционным, более подробно остановимся на возможных механизмах этих изменений. Природа лежащего в их основе явления представляется пока недостаточно ясной. В соответствии с литературными данными можно предположить, что анализируемый показатель в некоторой степени связан с медленными электрическими процессами (уровнем постоянного потенциала), которые, отражая активность нейронного ансамбля в целом, в свою очередь модулируют активность быстрых элементов системы, т.е. отражают звено регуляции, соединяющее энергетические и ионные механизмы с нейронной активностью (Н.А. Аладжа-лова, 1979; В.Ф. Фокин, Н.В. Пономарева, 1994). При этом быстрые и медленные процессы, развивающиеся на нейронных мембранах, зависят от всего комплекса биохимических процессов, влияющих на регуляцию мембранного потенциала нервных клеток.
Наши данные показывают, что усиленная неспецифическая афферентация при восприятии новых, неожиданных сигналов приводит к генерализованной позитивации биопотенциалов мозга в сочетании с их локальной негативацией в проекционных отделах, тогда как при вовлечении в процесс анализа информации кортикофугальных регулирующих влияний нега-тивация электрокорковой активности отмечается во всех исследованных областях. Корреляция между усилением корковой активности и негативацией биопотенциалов в процессе когнитивной деятельности показана в исследовании М. Lang с соавторами (М. Lang et al., 1989). Это может быть отражением
258
различного соотношения степени возбуждения поверхностных и глубоких слоев коры (апикальных дендритов и тел нейронов) (Н.А. Аладжалова, 1979). Следует отметить, что, в соответствии с литературными данными, возрастание импульсной активности нейронов коры коррелирует с доминированием в электрограмме коры высокочастотных компонентов, тогда как при возбуждении апикальных дендритов преобладают медленноволновые составляющие. С учетом этих сведений получают дополнительное объяснение и наши данные об увеличении удельного веса высокочастотных ритмов в паттерне активности, связанной со стимулом, особенно при повышении его сигнальной значимости (Л.А. Рожкова, 1997).
Выявленная региональная специфика амплитудных значений РД (максимум негативации в левой лобной зоне при восприятии релевантных стимулов), по-видимому, в большей степени отражает их связь с мембранными потенциалами данной популяции нейронов, чем с метаболическими факторами (В.Ф. Фокин, Н.В. Пономарева, 1994). Увеличение роли перед-неассоциативных отделов коры в процессах восприятия релевантной информации (в условиях нашего эксперимента - при счете стимулов) находит отражение в усилении негативации биопотенциалов этих структур при реакции на значимые стимулы, что, в соответствии с многочисленными литературными и собственными данными, рассматривается как повышение уровня корковой активации (Э.С. Ополинский, Л.А. Рожкова, 1991; Р. Наатанен, 1998; R. Naatanen, 1986). В этой связи следует отметить, что одним из преимущественных аппаратов медленной управляющей системы рассматривается лимбико-ги-поталамический комплекс, тогда как с деятельностью быстродействующего аппарата мозга связана в основном ретикулярная система (Н.А. Аладжалова, 1979); характер, соотношение степени их вовлечения, взаимодействия при осуществлении процессов регуляции, очевидно, и находят отражение в различных электрофизиологических параметрах.
Выявленные динамические сдвиги уровня поляризации биопотенциалов мозга при повышении значимости воспринимаемой информации, по-видимому, отражают деятельность регуляторных механизмов мозга, обеспечивающих большую или меньшую степень вовлечения в процесс восприятия тех или иных функциональных систем. Дальнейшее Исследование этой характеристики ЭА в норме и при различных нарушениях вовлечения в процесс восприятия спе-
259
цифических и неспецифических систем, вероятно, может способствовать более точному пониманию функционального смысла и природы данного показателя.
Таким образом, анализ особенностей возникновения и уга-шения ОР дает возможность оценить уровень развития нейрофизиологических механизмов (в частности, процессов активации, регуляции, следообразования), обеспечивающих структурно-функциональную реорганизацию в ЦНС ребенка, адекватную осуществляемому виду деятельности, что рассматривается как основной системный механизм обучения и памяти. В комплексе с клиническим и психолого-педагогическим обследованием данный метод представляется наиболее значимым при диагностическом исследовании детей с трудностями обучения различной этиологии, а это может расширить возможности коррекционных мероприятий с учетом ведущего фактора в структуре дефекта.
7.3. Нейрофизиологические критерии диагностики
нарушений познавательной деятельности у детей
с задержкой психического развития
В данном разделе мы попытались показать информативность комплексного изучения характеристик "фоновой" ЭЭГ и различных компонентов ориентировочной реакции (ОР) на сенсорные стимулы для оценки степени сформированности или сохранности нейрофизиологических механизмов внимания и памяти у детей с ЗПР младшего, среднего и старшего школьного возраста. Представлено соотношение особенностей ОР у детей с ЗПР двух основных подгрупп, ведущим фактором в структуре дефекта одной из которых по поведенческим характеристикам является, главным образом, дефицит внимания (ЗПР/ДВ), а другой - недостаточность преимущественно мнестических процессов (ЗПР/ДП).
7.3.1. Особенности ЭЭГ в состоянии спокойного бодрствования
Нарушения развития познавательной деятельности, в частности, дефицит произвольного внимания, трудности запечат-ления и воспроизведения информации могут быть обусловлены недостаточной зрелостью, особенностями функционального состояния кортикальных и субкортикальных структур моз-
260
га. Это закономерно проявляется в уровне сформированности ритмической активности (оцениваемом по характеристикам спектральной плотности мощности основных ритмов ЭЭГ и по данным визуального анализа), а также в особенностях ее зонального распределения, межполушарной асимметрии, наличии знаков дисфункции ЦНС, выявляемых при регистрации ЭЭГ в покое и при функциональных нагрузках. Характеристики "фоновой" ЭЭГ, как уже отмечалось, в меньшей степени, чем параметры ОР, отражают специфику деятельности мозга детей с ЗПР. Рассматриваемые в парадигме традиционных клинических исследований, они оказываются в ряде случаев недостаточно информативными для диагностики ЗПР. Однако в совокупности с показателями ОР эти данные способствуют более глубокому пониманию причин выявляемых отклонений в развитии ребенка.
ЭЭГ подгруппы детей с ЗПР в целом характеризуется такими особенностями, которые в контексте с результатами анализа ОР могут рассматриваться как признаки структурно-функциональной недостаточности различных структур головного мозга. Так, спектральный анализ мощности основных ритмов фоновой ЭЭГ показал, что у детей с ЗПР/ДВ младшего школьного возраста отмечается большая по сравнению с нормой мощность ритмов тета-диапазо-нов (тета 1 и тета 2 в затылочной области коры; тета 2 - в центральной корковой зоне).
Спектральная мощность ритмики альфа 1 и альфа 2 диапазонов характеризуется значительной межиндивидуальной вариабельностью, что делает нецелесообразным сопоставление средних групповых значений спектральной мощности. На рис. 7.2 представлена частота встречаемости различных значений спектральной мощности ритма альфа 1. Обращает на себя внимание то, что в норме распределение значений мощности в трех избранных диапазонах (0-10; 10-20; 20 усл. ед. и более) приближается к нормальному — на центральный интервал приходится наибольшее число случаев. У детей с ЗПР/ДВ младшего школьного возраста чаще, чем в норме, встречаются случаи с низкими значениями мощности ритма альфа 1. Ритм диапазона альфа 2 встречается почти в два раза реже, чем ритм альфа 1, что характерно и для нормы и обусловлено более поздним созреванием генераторов высокочастотных составляющих альфа-активности.
261

Рис. 7.2. Частота встречаемости различных значений спектральной мощности альфа 1- и альфа 2-ритмов в задних отделах коры левого полушария у детей младшего (светлые столбцы), среднего (серые столбцы) школьного и подросткового (темные столбцы) возраста в норме и при ЗПР. По оси абсцисс - диапазоны значений мощности альфа-ритма в усл. ед. По оси ординат - процент случаев
У детей с ЗПР/ДВ в возрасте 9,5-12 лет чаще, чем в норме встречаются случаи с низкими значениями мощности альфа 2 ритма. В старшем школьном возрасте у 55% детей с ЗПР, напротив, обнаруживается гиперсинхронный альфа 2 ритм, в то время как у нормально развивающихся сверстников альфа 2 ритм повышенной мощности регистрируется гораздо реже
262
(в 30% случаев). Наряду с этим и в отличие от нормы более чем у 30% подростков с ЗПР еще существенно выражена ритмика альфа 1 диапазона, а в 10% случаев он имеет гиперсинхронный характер; ритмика альфа 2 диапазона, напротив, более чем в 20% случаев выражена слабо.
Визуальный анализ выраженности различных признаков дисфункции структур мозга показывает, что у детей с ЗПР/ДВ чаще, чем в норме, отмечается нерегулярность, дезорганизация альфа-ритма. Наличие медленных волн в задних отделах коры, указывающее на заинтересованность стволовых структур мозга (по одним данным — преимущественно нижнестволовых, по другим - диэнцефальных (Л.Р. Зенков, 1996)), не является специфичной особенностью функционального состояния мозга детей с ЗПР 6,5-9 лет, т.к. они достаточно часто регистрируются и в норме. Билатерально-синхронная пароксизмальная активность в норме отмечается в единичных случаях как в покое, так и при функциональных нагрузках, в то время как у детей с ЗПР/ДВ в обеих ситуациях она встречается в 30% случаев.
В целом обнаруженная у детей с ЗПР/ДВ большая сравнительно с нормой выраженность ритмики тета 2 диапазона, более частая встречаемость нерегулярности и малой мощности альфа-активности могут указывать на незрелость корково-подкорковых взаимодействий (Н.Н. Зислина и др., 1972; Н.В. Дубровинская, 1985; И.П. Лукашевич и др., 1998). Эти признаки незрелости, хотя и в меньшей степени, проявляются у детей с ЗПР/ДВ и в старшем школьном возрасте. Повышенная, избыточная сравнительно с нормой выраженность медленной активности тета-диапазона в сочетании с недостаточной представленностью высокочастотных составляющих ЭА (особенно бета-диапазона) обнаружена у детей с ДВ в исследовании Дж. Любар (J.F. Lubar, 1993). Такое соотношение, проявляющееся в существенном превышении мощности тета-активности над бета, отмечается как в покое, так и, в особенности, при выполнении заданий, что рассматривается автором как лучший показатель, отражающий корковую гипоактивацию.
При рассмотрении особенностей ЭЭГ подгруппы ЗПР/ДП обнаруживаются более существенные отличия от нормы на всех исследованных этапах онтогенеза, особенно демонстративные при сравнении характера распределения индивидуальных значений СПМ основных ритмов ЭЭГ. Наиболее выраженная
263
возрастная динамика отмечается для ритмов альфа 1 и альфа 2 диапазонов. Как следует из рис. 7.2, в норме с возрастом увеличивается частота встречаемости альфа 1 ритма с низкой спектральной мощностью (до 5 условных единиц) и альфа 2 ритма более высокой СПМ. В отличие от нормы среди детей с ЗПР и дефицитом памяти младшего, среднего и даже старшего школьного возраста отмечается высокая частота встречаемости гиперсинхронного альфа 1 ритма (67, 45 и 25% соответственно). При этом ритмика альфа 2 диапазона в большинстве случаев имеет низкую мощность за исключением старших школьников, у которых в 63% случае, т.е. вдвое чаще, чем в норме, регистрировали гиперсинхронный альфа 2 ритм (что отмечалось и в группе ЗПР/ДВ). Гиперсинхрония альфа-активности в каудаль-ных отделах у детей с трудностями обучения отмечается и в других исследованиях (И.П. Лукашевич и др., 1994, 1998; J. Becker et al, 1987). В работах И.П. Лукашевич с соавторами (1994, 1998) эти данные рассматриваются как показатель "заинтересованности", дисфункции нижних отделов ствола мозга, которая, как полагают, обусловлена угнетением возбуждающих влияний со стороны ретикулярной формации ствола мозга и усилением синхронизирующих воздействий системы Моруцци. Такие особенности корковой ритмики в период полового созревания могут быть обусловлены гиперактивностью гипоталами-ческих структур (Д.А. Фарбер, Н.В. Дубровинская, 1989), которая в большей степени проявляется у детей с ЗПР в связи с присущей им недостаточностью коркового контроля.
Отмечающаяся у детей с ЗПР/ДП несформированность ритмической активности альфа-диапазона (рассматриваемая как отражение слабого развития циклических процессов в ЦНС и коррелирующая с низким уровнем ее пластичности) может приводить к снижению эффективности прогностической деятельности. Это предположение подкрепляется результатами исследований Е.А. Громовой (1980), в которых показано, что для детей с ЗПР с нерегулярным, редуцированным альфа-ритмом характерно снижение уровня экскреции биогенных аминов (серотонина), оказывающих стимулирующее влияние на следовые процессы. Гиперсинхрония альфа-ритма, обнаруженная у части детей с ЗПР, по-видимому, также может обусловливать недостаточную реактивность коры головного мозга на новую или маловероятную информацию, а следовательно, и приводить к ухудшению процессов следообразо-вания, формирования НМС.
264
Среди детей с ЗПР, для которых характерна недостаточность процессов запечатления и воспроизведения информации, и в младшем, и в старшем школьном возрасте отмечается высокая частота встречаемости в ЭЭГ таких знаков дисфункции ЦНС, как пароксизмальные билатеральные вспышки медленноволновой или гетерочастотной активности стволового или диэнцефального генеза при гипервентиляционных пробах (в 62% случаев), замедленные колебания в задних отделах коры мозга (25% случаев). Нерегулярность альфа-ритмики отмечается у 56% обследованных старшеклассников.
Следовательно, у детей с ЗПР и дефектами памяти на всех исследованных этапах возрастного развития обнаруживаются признаки недостаточности нижнестволовых и диэнцефальных структур головного мозга. Независимая от возраста устойчивость выраженности ЭЭГ-знаков, указывающих на аномальное состояние диэнцефальных структур мозга у детей с ДП, вероятно, свидетельствует об органической природе этих изменений и скорее о неполноценности, чем о незрелости связанных с этими структурами процессов, что служит дополнительным подтверждением предположения о нарушении у этих детей преимущественно нейрофизиологических механизмов памяти. Разнообразные нарушения мнестических процессов при повреждении лимбико-диэнцефальных и мезэнцефальных структур головного мозга широко описаны в литературе (А.Р. Лурия, 1974; Н.П. Бехтерева, 1985; Н.К. Корсакова, Л.И. Московичюте, 1985).
7.3.2. Характеристики ориентировочной реакции
А. Условия непроизвольного внимания. Основной отличительной особенностью ОР на индифферентные стимулы у обеих групп детей с ЗПР младшего школьного возраста является существенно меньшая по сравнению с нормой выраженность вегетативного (КГР) и электрокоркового (РД) компонентов ОР, причем наибольшие отличия присущи детям с ЗПР/ДВ.
Следует отметить, что у младших школьников как в норме, так и при ЗПР реакция активации, проявляющаяся в виде Десинхронизации электроактивности мозга (корковый компонент ОР), характеризуется неустойчивостью: после предъявления первых трех стимулов она выявляется нерегулярно, что Делает нецелесообразным анализ динамики ее угашения.
265


Рис. 7.3. Особенности реакции десинхронизации электроактивности затылочной и центральной зон коры левого полушария в условиях непроизвольного и произвольного внимания у двух групп детей с ЗПР: с дефицитом внимания (ЗПР/ДВ) и дефицитом памяти (ЗПР/ДП) в сопоставлении с нормой в различные возрастные периоды. По оси абсцисс -возраст; по оси ординат - длительность реакции десинхронизации в мс
Длительность реакции активации на индифферентные зрительные стимулы в проекционной (затылочной) зоне коры у младших школьников с ЗПР/ДВ почти в полтора раза меньше, чем у нормально развивающихся детей. Напротив, в среднем и старшем школьном возрасте этот показатель превышает присущие норме значения (на 35 и 20% соответственно) (рис. 7.3). Такая "избыточная" активация коры головного мозга может быть причиной обнаруживаемой в структуре синдрома дефицита внимания повышенной чувствительности к внешним раздражителям.
266
В отличие от детей с ЗПР/ДВ при ЗПР/ДП возрастные изменения ДРД как затылочной, так и центральной зон коры на иррелевантные стимулы несущественны, в связи с чем и в старшем школьном возрасте ее значения остаются меньше нормативных (см. рис. 7.3).
Анализ соотношения ДРД в проекционной и ассоциативной коре в условиях непроизвольного внимания показывает, что у всех групп детей продолжительность реакции активации в центральной зоне меньше, чем в затылочной. Однако, у детей с ЗПР/ДВ младшего возраста и у всех детей с ЗПР/ДП эти различия выражены значительно меньше, чем в норме (см. рис. 7.3).
Усиление связанной со стимулами реакции десинхронизации у детей с ЗПР/ДВ коррелирует с увеличением частоты встречаемости КГР в состоянии покоя (тогда как в норме этот показатель с возрастом уменьшается): в среднем школьном возрасте КГР регистрируется почти в два раза чаще, а в старшем - в четыре раза чаще, чем в младшем (когда она отмечалась лишь в 15% случаев) и на 15% чаще, чем в норме.
При ЗПР/ДП закономерной возрастной динамики КГР не наблюдается.
Б. Условия произвольного внимания. Сдвиги ДРД при повышении уровня внимания у детей с ЗПР/ДВ аналогичны выявленным в норме : уменьшение ДРД в затылочной коре, возрастание - в центральной. Исключение составляют младшие школьники, для которых, в отличие от нормы, характерно увеличение продолжительности реакции активации в обеих зонах коры, но более выраженное в центральной области (рис. 7.4).
У детей с дефицитом памяти (ЗПР/ДП) направление сдвигов ДРД затылочной коры на релевантные стимулы по сравнению с таковой на индифферентные совпадает с наблюдаемыми в норме. Однако в центральной области коры эти изменения, в отличие от нормы, несущественны: в младшей и старшей возрастных группах отмечается крайне слабое увеличение ДРД (всего на 8,8 и 4% соответственно от исходного уровня), а в среднем школьном возрасте ее значения даже уменьшаются на 22% (см. рис. 7.4).
Таким образом, повышение значимости информации (путем привлечения внимания к стимулам с помощью речевой инструкции), способствующее усилению регулирующих влияний со стороны лобных отделов коры, приводит к адекватному компенсаторному увеличению длительности реакции активации только у детей с ЗПР/ДВ.
267


Рис. 7.4. Влияние уровня внимания на длительность реакции активации в затылочной и центральной зонах коры левого полушария у двух групп детей с ЗПР: с дефицитом внимания (ЗПР/ДВ) и дефицитом памяти (ЗПР/ДП) в сопоставлении с нормой в различные возрастные периоды. Серые столбцы - непроизвольное внимание (НПВ); темные - произвольное внимание (ПВ). Остальные обозначения - как на рис. 7.3
Отсутствие выраженной реорганизации степени вовлечения различных областей коры при повышении значимости воспринимаемых стимулов, характерное для группы с ДП, может служить, на наш взгляд, информативным показателем для оценки нейробиологических предпосылок развития некоторых психических функций при аномалиях онтогенеза.
Вместе с тем важно отметить тот факт, что для детей с ЗПР/ДВ каждого из исследованных возрастных интервалов характерна менее продолжительная, чем в норме, реакция активации центральных областей коры на релевантные сигналы, хотя ее длительность и увеличивается в онтогенезе. Эти различия существенны как в младшем, так и в старшем школьном возрасте, что может свидетельствовать о сохране-
268
нии структурно-функциональной недостаточности переднеас-социативных отделов коры, неполноценности их регулирующих влияний по сравнению с нормой (С.A. Riccio et al., 1993), возможно, вследствие незрелости этих отделов мозга и их связей с подкорковыми образованиями. Это означает, что не обеспечивается тот уровень активации мозга, который необходим для оптимального протекания процессов переработки сенсорной информации и решения познавательных задач. Полученные результаты согласуются с имеющимися в литературе данными о сниженном уровне корковой активации у детей с дефицитом внимания, сочетающимся с гиперактивностью. При этом одни авторы рассматривают этот факт как отражение ослабления стволовых активирующих влияний (J.H. Satterfield, M.E. Dawson, 1971; D. Cantwell, 1977), тогда как другие - как следствие дисфункции системы переднего мозга (лимбические структуры и лобная кора), обусловливающей слабую модуляцию активности ретикулярной формации ствола (Е. Grunewald-Zuberbier et al., 1977; R.H. Rosenthal, T.W. Allen, 1978; R.A. Dykman, P.T. Ackerman, 1993).
У детей группы ЗПР/ДВ младшего возраста низкий уровень корковой активации сочетается с меньшей, чем в норме, выраженностью спонтанных КГР в покое, меньшей реактивностью КГР в условиях непроизвольного внимания, приближающейся к нормативным величинам лишь в ситуации произвольного внимания. Эти данные можно рассматривать как отражение некоторой недостаточности эмоциогенных лимбико-диэнцефальных структур, компенсируемой регулирующими нисходящими корковыми влияниями. Незрелость эмоционально-волевой сферы - одна из основных психологических и клинических характеристик детей с ЗПР различного генеза. В ряде случаев она является основным симптомом ЗПР и может сглаживаться с возрастом по мере формирования передних отделов коры и их связей с диэнцефальными структурами (М.С. Певзнер, 1972; Актуальные проблемы..., 1982).
У детей с недостаточностью мнестических процессов возрастная динамика ДРД затылочной и центральной зон коры на релевантные стимулы выражена еще в меньшей степени, чем при ЗПР/ДВ. В связи с этим при ЗПР/ДП даже в старшем школьном возрасте ДРД в затылочной коре практически не отличается от своих значений у младших нормально развивающихся школьников, а в центральной области коры не достигает таковых.
269
Особенно важным, на наш взгляд, диагностическим критерием является характер соотношения степени вовлечения в ОР проекционной и ассоциативной зон коры в условиях направленного внимания по сравнению с таковым при восприятии индифферентных стимулов.
При ЗПР/ДВ во всех возрастных диапазонах, как и у нормально развивающихся школьников, длительность реакции активации в центральной коре в условиях произвольного внимания становится больше, чем в затылочной, хотя у детей с ЗПР/ДВ эти различия выражены гораздо слабее, чем в норме, особенно в младшем школьном возрасте. Напомним, что в ситуации ненаправленного внимания более интенсивная реакция активации характерна для проекционной (затылочной) коры (см. рис. 7.3, 7.4).
При ЗПР/ДП ни в одной возрастной группе, в отличие от нормы и детей с дефицитом внимания, при предъявлении релевантной информации значения ДРД в центральной корковой зоне не превышали таковые в затылочной коре (см. рис. 7.3).
Кроме оценки средних значений ДРД на первые три стимула ряда информативным для диагностики ЗПР/ДП у детей среднего и старшего школьного возраста является анализ динамики изменений ДРД при угашении ОР (анализ динамики ДРД у младших школьников затруднен ввиду нерегулярности появления реакции десинхронизации). У детей с нарушениями памяти отмечается менее выраженный, чем в норме, декремент ДРД центральной области коры при предъявлении значимых стимулов (рис. 7.5). Это может быть обусловлено слабостью ассоциативных механизмов мозга, процессов следообразования, лежащих в основе уга-шения ОР, "негативного научения" (Е.Н. Соколов, 1969; Б.И. Котляр, 1989).
В целом можно сделать вывод, что нарушения памяти у детей с ЗПР коррелируют как с особенностями ЭЭГ в состоянии покоя, так и, что наиболее важно, со спецификой реорганизации биоэлектрической активности мозга при повышении значимости поступающей информации (слабое вовлечение центральных областей коры в анализ релевантных стимулов).
Совокупность полученных электрофизиологических данных позволяет охарактеризовать причины слабости процессов регуляции, выявляемой по поведенческим характеристикам, которая в разном возрасте может иметь неодинаковую нейрофизиологическую основу.
270

Рис. 7.5. Динамика угашения реакции десинхронизации в центральной области коры левого полушария на релевантные стимулы у детей среднего школьного (пунктир) и подросткового (сплошная линия) возраста в норме и при разных вариантах ЗПР. По оси абсцисс -порядковые номера стимулов; по оси ординат - длительность реакции десинхронизации в мс
271
У младших школьников нарушения внимания могут быть связаны с гипофункцией активирующей ретикулярной формации ствола мозга, незрелостью корково-подкорко-вых взаимодействий, обусловленной недостаточной сфор-мированностью лобных отделов коры головного мозга и системы их регулирующих влияний. Эта незрелость в первую очередь может быть причиной трудностей привлечения внимания к новой информации, неустойчивости внимания при необходимости выполнения длинного ряда операций (Актуальные проблемы..., 1982; Дети с задержкой..., 1984; Диагностика и..., 1988; Л.И. Переслени, 1984; Ю.Д. Кропо-тов и др., 1999).
Гипофункция восходящей активирующей системы, обусловленная незрелостью корково-подкорковых взаимодействий у детей младшей возрастной группы, сменяется ее гиперфункцией. В связи с этим дефицит внимания, обнаруживаемый у части детей с ЗПР старшего возраста, приобретает другой функциональный смысл, чем у младших школьников. Это не столько низкая устойчивость внимания, сколько недостаточная селективность, трудности его переключения и распределения. Неспособность игнорировать иррелевант-ные стимулы может быть связана с дисфункцией мезолим-бической дофаминэргической системы (ДАС), опосредующей развитие латентного торможения (Е.А. Громова, 1980; J. Feldon, I. Werner, 1988; С.А. Riccio et al., 1993; A. Talay-Ongan, 1998). Имеются также данные о том, что ДАС наиболее интенсивно иннервирует лимбическую и ассоциативную кору, причем картина иннервации зон и слоев коры свидетельствует о том, что эта система может сильнее влиять на активность корково-корковых, чем таламо-корко-вых связей; следовательно, ДАС участвует скорее в высших интегративных процессах, чем в сенсорном анализе (S.L. Foot, J.H. Morrison, 1987).
Недостаточная сформированность произвольного внимания, эмоционально-волевой сферы при относительно сохранных познавательных процессах на всех этапах возрастного развития требует направленной педагогической коррекции, что может способствовать преодолению трудностей обучения.
Особенности ОР могут служить обоснованием имеющихся в литературе данных (Ф.Н. Гоноболин, 1972; S.S. Zental, 1993) о том, что для привлечения внимания детей младшего школь-
272
ного возраста целесообразно использовать в системе обучения такие факторы, как яркость, громкость и другие физические параметры объектов восприятия, непосредственно воздействующих на органы чувств. В то же время избыточность непосредственно воздействующих раздражителей для детей с ЗПР среднего и старшего школьного возраста в соответствии с обнаруженными нами фактами (избыточная сравнительно с нормой активация коры) может ухудшать усвоение знаний и формирование представлений.
Совокупность полученных данных позволяет сделать вывод о том, что для повышения эффективности обучения детей с нарушениями регуляции произвольных форм деятельности как в младшем, так и в старшем школьном возрасте необходимо включение в систему коррекционных мероприятий разнообразных приемов, усиливающих интерес ученика к поступающей из внешней среды информации, повышающих ее значимость для ребенка.
При обучении детей с ЗПР, у которых ведущим фактором в структуре дефекта является недостаточность познавательных процессов (памяти, восприятия, мышления), необходимо использование иных коррекционных воздействий, чем для детей с нарушениями регуляции произвольных форм деятельности. Можно полагать, что все приемы, способствующие лучшему запечатлению воспринятого материала, направленного, разнообразного воспроизведения пройденного, рекомендуемые для обучения умственно отсталых школьников, целесообразно использовать и в системе обучения детей с ЗПР. Это в первую очередь относится к тем детям, у которых ведущим фактором в структуре нарушений познавательной деятельности являются нарушения памяти, восприятия, мышления. Все, что способствует установлению взаимосвязи между событиями внешнего мира, появлению интереса, адекватной мотивации, может быть полезно для развития ассоциативных процессов и способствовать повышению эффективности познавательной деятельности.
При сопоставлении поведенческих характеристик внимания и памяти с показателями ОР и с особенностями ЭЭГ у одних и тех же детей появляется дополнительная возможность подойти к более глубокому пониманию механизмов, лежащих в основе тех или иных проявлений недостаточности когнитивной деятельности.
273
7.4. Нейрофизиологические критерии оценки уровня развития механизмов когнитивной деятельности детей с речевой патологией
В данном разделе представлены результаты анализа характеристик ЭЭГ в состоянии покоя и компонентов ОР на индифферентные и значимые стимулы у двух групп детей с ОНР младшего школьного возраста (7-9,5 лет) - с нарушениями преимущественно процессов внимания (ОНР/ДВ) или памяти (ОНР/ДП), которые, как и дети с ЗПР, были дифференцированы в соответствии с данными психологического обследования и педагогическими характеристиками.
7.4.1. Особенности ЭЭГ в состоянии спокойного бодрствования
Несмотря на значительный полиморфизм состава детей с речевой патологией одной из существенных отличительных особенностей ЭЭГ группы детей с ОНР в целом является нерегулярность ритмики альфа-диапазона (более 50% случаев), причем у детей с преобладанием нарушений памяти это отмечается гораздо чаще, чем у детей с нарушениями внимания (42 и 5% случаев соответственно). У них более часто, чем в норме, встречается альфа 2 ритм с низкой (менее 10 мкв/Гц) СПМ (58 и 14,5% случаев соответственно).
Ритмика альфа 1 диапазона у детей с нарушениями памяти также отличается несформированностью и регистрируется лишь в 25% случаев, тогда как в норме и у детей с ОНР и нарушениями внимания он представлен в 64 и 60% случаев соответственно. Следует также отметить, что у детей с нарушениями памяти чаще, чем в норме и при дефиците внимания, отмечалось неправильное зональное распределение корковой ритмики. Характерный для нормальной ЭЭГ фокус альфа-ритма в задних областях коры отмечается у этих детей лишь в 25% случаев для альфа 1 и в 50% для альфа 2; помимо этого отмечается его доминирование в височных, центральных и лобных отделах коры.
Анализ уровня развития ЭА коры мозга (по степени сформированное™ альфа-активности и характеристикам реакции следования ритму световых мельканий в диапазоне частот 4-20 Гц) показал, что несоответствие этого показателя возрастным нормативам у детей с ДП отмечалось в 50% случаев, тогда как в группе с ДВ - только в 3%.
274
Существенной особенностью структурно-функциональной организации мозга детей с ОНР является и выявленная в результате спектрального анализа специфика асимметрии корковой ритмики. Наибольшие отличия обнаружены при анализе асимметрии высокочастотной активности бета 1и бета 2 диапазонов, которая в норме в большинстве случаев (63% для бета 1 и 88% для бета 2) носила левосторонний характер (преобладание СПМ в левополушарных корковых зонах, чаще в передне- и задневисочных, а при ОНР, особенно с выраженной мнестической недостаточностью, имела противоположную направленность - в 100 и 75% случаев отмечалось преобладание СПМ ритмики бета 1 и бета 2 диапазонов соответственно в корковых областях правого полушария, причем наибольшие межполушарные различия в 50% случаев были характерны для задневисочных отделов коры. Дети с ОНР и дефицитом внимания отличались от нормы по этому показателю в меньшей степени (левостороннее доминирование бета 1-й бета 2-ритмов выявлено в 40-50% случаев).
Выявляемое своеобразие межполушарной асимметрии ритмической активности бета-диапазона (большая мощность бета-ритма не в левом, как в норме, а в правом полушарии) свидетельствует о специфике структурно-функциональной организации мозга детей с ОНР. Это представляется закономерным, если иметь в виду сведения об уменьшении мощности бета-активности у детей с дислексией по сравнению с нормой (G. Fein et al., 1986), а также о положительной корреляции выраженности бета-ритма в левом полушарии с успешностью выполнения вербальных и пространственных задач (D. Giannitrapani, 1985).
Низкочастотная ритмика тета 1 и тета 2 диапазонов в норме и в группе с ОНР в целом чаще (88 и 70% случаев соответственно) доминировала в правополушарных областях, причем наибольшая асимметрия отмечалась в задневисочных корковых зонах. Однако в группе ОНР с нарушениями памяти чаще, чем в других выборках, встречался противоположный характер асимметрии тета-ритмов - левостороннее доминирование (50% случаев).
Ритмика альфа-диапазона характеризовалась менее отчетливой асимметрией с тенденцией к преобладанию в задних (затылочных, височных) отделах коры правого полушария у детей в норме и с ОНР/ДВ. Однако наряду с этим в 57% случаев в норме отмечалось левополушарное домини-
275
рование альфа-ритма в передневисочных и центральных корковых зонах и только в 14% - в затылочной коре. В группе ОНР/ДП помимо характерного для нормы доминирования альфа-ритма в задних отделах коры правого полушария в 50% случаев отмечалось его преобладание в лево-полушарной затылочной области.
Наряду с отмеченными общемозговыми нарушениями (проявляющимися в несформированности альфа-ритма, особенностях его зонального распределения, межполушарной асимметрии ритмических составляющих ЭЭГ) у детей с ОНР выявляются и очаговые изменения корковой ритмики, указывающие на повреждение определенных мозговых структур. Такие изменения, проявляющиеся в наличии патологической или пароксизмальной активности, отмечались у 30% детей с ОНР. Важно подчеркнуть, что у детей с недостаточностью мнестических процессов локальные изменения отмечаются гораздо чаще, чем у детей с ДВ (61 и 16% случаев, соответственно). При этом в 40% случаев в группе ОНР/ДП они были зарегистрированы в правом полушарии (преимущественно в височно-теменно-затылочных областях) и в 21% - в левополушарных структурах.
Билатерально-синхронные вспышки медленноволновой или гетерочастотной активности пароксизмального характера в фоновой ЭЭГ отмечались почти у 30% детей с речевой патологией. При этом в группе с ДП чаще (в 40%) отмечалась па-роксизмальная активность диэнцефального генеза; кроме того, у 68% этих детей в передних и центральных отделах наблюдалось выраженное преобладание тета-активности над альфа-ритмом, что ассоциируется с дисфункцией преимущественно верхнестволовых структур мозга (И.П. Лукашевич и др., 1994, 1998). Важно также отметить, что пароксизмальные изменения ЭА лобно-базального генеза (разряды острых волн бета-диапазона в лобных, центральных и височных отделах, чаще с левополушарным акцентом) отмечались только в группе с ДП (16%). Указания на общемозговые нарушения иногда в комбинации с очаговыми и пароксизмальными проявлениями обнаружены при ЭЭГ-обследовании детей с недоразвитием речи и в других исследованиях (Нарушение речи..., 1972). Большая выраженность описанных изменений ЭА у детей с трудностями запечатления и воспроизведения информации указывает, с одной стороны, на большую тяжесть патологического процесса при ДП, нарушающего нормальный ход морфофункцио-нального развития мозга, а с другой стороны на тесную связь
276
этих изменений с нарушениями механизмов памяти, что сочеталось и с большими нарушениями в речевой деятельности.
Таким образом, выявленные особенности фоновой биоэлектрической активности свидетельствуют о существенных отклонениях в структурно-функциональной организации мозга детей с речевой патологией, характер и степень выраженности которых коррелирует с характером нарушений их познавательной деятельности.
7.4.2. Особенности ориентировочной реакции
А. Условия непроизвольного внимания. Сравнительный анализ характеристик компонента РЗОО у разных групп детей выявил существенные межгрупповые различия прежде всего его амплитудных значений (главный эффект фактора (ГЭФ) Объект: р < 0,05, наиболее выраженный в Тр). При этом для группы ОНР/ДВ характерна меньшая, чем в норме, величина РЗОО в каудальных отделах (в особенности в теменной коре и задневисочной зоне левого полушария), тогда как у детей с ОНР/ДП его амплитуда (особенно в затылочных зонах правого полушария), напротив, превышает таковую в норме (р < 0,05; рис. 7.6, А). Выраженные межгрупповые различия выявляются также в лобных отделах, где сдвиги потенциалов имеют противоположную направленность: негативация в норме и позитивация при ОНР, причем наиболее закономерные отличия от нормы отмечены для группы ОНР/ДП (р < 0,10). Следует обратить внимание на то, что наибольшие различия между двумя группами с ОНР выявляются в каудальных отделах (О, Тр), особенно левого полушария, где амплитуда РЗОО у детей с ОНР/ДВ меньше, чем в группе ОНР/ДП (р < 0,05).
Существенные межгрупповые различия характерны и для ПЛ РЗОО (ГЭФ Объект: р < 0,0004) и наиболее выражены в теменной и задневисочной зонах обоих полушарий; наибольшие значения ПЛ выявлены в норме, а наименьшие - в группе ОНР/ДВ (р < 0,05). У детей группы ОНР/ДВ отмечается также более короткая по сравнению с нормой и группой ОНР/ДП ПЛ РЗОО билатерально в затылочных и в левой передневисоч-ной зонах (р < 0,05; рис. 7.6, Б).
Отличительной особенностью РЗОО при ОНР является также меньшая, чем в норме, выраженность межзональных различий его амплитуды. Так, если в норме амплитудный максимум РЗОО отмечается в теменных отделах обоих полушарий,
277



Рис. 7.6. Особенности электрокорковых компонентов ОР в различных областях коры левого (L) и правого (R) полушарий мозга у двух групп детей с ОНР (с дефицитом внимания - ОНР/ДВ и дефицитом памяти -ОНР/ДП) в сопоставлении с нормой в условиях непроизвольного внимания. Среднегрупповые значения: А - амплитуды компонента РЗОО (мкВ/мс); Б - пиковой латснтности РЗОО (в мс); В - величины сдвига ЭА в период РД (мкВ/мс)
но наиболее четко - в левом (что проявляется в преобладании его амплитуды в теменной коре по сравнению со всеми другими областями - р < 0,03), то в обеих группах с ОНР это выражено в меньшей степени: амплитуда РЗОО в теменной коре превышает таковую лишь в височных зонах (р < 0,05); в группе ОНР/ДВ в обоих полушариях отмечено ее преобладание и сравнительно с лобной корой (р < 0,05 - рис. 7.6, А).
278
Иные, чем в норме, межрегиональные соотношения присущи и ПЛ РЗОО. Напомним, что в норме максимальная ПЛ отмечена в задневисочной коре обоих полушарий при относительно наименьших ее значениях в симметричных центральных зонах, а также в теменной, затылочной и лобной коре левого полушария (р < 0,05). У детей с ОНР/ДВ наиболее корот-колатентный РЗОО зарегистрирован в теменной коре правого полушария (р < 0,05 в сравнении с зонами С, Та и Тр), а в левом полушарии межзональные различия ПЛ не проявились. У детей с ОНР/ДП, напротив, в левом полушарии выявлена достоверно меньшая ПЛ РЗОО в теменной коре сравнительно с затылочной и задневисочной зонами (р < 0,05; рис. 7.6, Б).
Таким образом, свойственная норме специфика участия теменных отделов коры (особенно левого полушария) в генерации РЗОО у детей с ОНР (в особенности сочетающимся с нарушениями преимущественно мнестических процессов) выражена меньше, а вовлечение лобной коры носит иной, противоположный характер.
Следует отметить также меньшую, чем в норме, межполу-шарную асимметрию амплитуды РЗОО у детей с ОНР: в обеих группах асимметрия проявилась лишь в виде тенденции к преобладанию РЗОО в левой затылочной зоне по сравнению с правой (р < 0,15).
Отличительной особенностью детей с ОНР является также более слабая, чем в норме, выраженность другого электрокоркового компонента ОР - реакции десинхронизации корковой ритмики, что проявляется в меньшей ее длительности в затылочной и в особенности в центральной областях коры (табл. 7.2).
Максимальные различия ДРД по сравнению с нормой характерны для группы детей с ОНР и нарушениями памяти, наиболее существенные при сопоставлении выраженности реакции десинхронизации в центральных корковых зонах (меньше на 195 мс).
Выявленные особенности длительности РД сочетались и со спецификой поляризации ЭА в период ее развития (суммарный эффект фактора Объект: р < 0,009, наиболее выраженный в С и F). Так, в группе ОНР/ДП обнаружена меньшая, чем в норме, позитивация ЭА в переднеассоциативных отделах (рис. 7.6, В). Но в наибольшей степени отличались от нормы по данному показателю дети группы ОНР/ДВ (р < 0,001): у них в большинстве корковых зон (кроме Та) вы-
279
Таблица 7.2
Особенности реакции десинхронизации (длительность, мс)
в затылочной (Os) и центральной (Cs) областях коры левого
полушария в условиях непроизвольного (1) и произвольного (2)
внимания у детей в норме и при ОНР (в целом и в подгруппах ОНР/ДВ и ОНР/ДП)
|
|
Os |
Cs | ||
|
|
1 |
2 |
1 |
2 |
|
Норма 680 |
525 465 |
665 | ||
|
ОНР 520 |
460 310 |
425 | ||
|
ОНР/ДВ 560 |
410 320 |
520 | ||
|
ОНР/ДП 510 |
530 270 |
290 | ||
явлена негативация ЭА, наиболее выраженная в левом полушарии, где различия сравнительно с нормой достоверны в теменной, центральной и лобной зонах (р < 0,05), а в правом полушарии — лишь в лобной коре (р < 0,002). При этом максимальная негативация в обоих полушариях отмечается во фронтальной коре (р < 0,05 сравнительно с височными зонами и левой затылочной; рис. 7.6, В). Аналогичные особенности отличают детей с ОНР/ДВ и от группы ОНР/ДП (ГЭФ Объект: р < 0,006; для F3 и F4: р < 0,03).
Следует также отметить, что у детей обеих групп с ОНР в отличие от нормы обнаружен выраженный градиент потенциалов в задневисочных зонах - от позитивности РЗОО к негативности в период РД. Представленные в данном разделе данные свидетельствуют о специфике структурно-функционального обеспечения даже такого сравнительно простого перцептивного акта как ОР на вспышку света у детей с нарушениями речевого развития. Группу детей с ОНР в целом отличают от нормы следующие особенности реализации ОР: отсутствие преимущественного (по амплитуде РЗОО) вовлечения теменной коры (особенно левого полушария); большая выраженность РЗОО в лобных отделах (в норме в этот период отмечена негативация ЭА); меньшая ДРД и в проекционных, и в ассоциативных зонах коры; меньшая позитивация ЭА в период развития РД, особенно в переднеассоциативных (лобных и центральных) отделах коры, а также в теменной области левого полушария.
Важными факторами, определяющими специфику реализации данного процесса, являются особенности развития и/или состояния механизмов внимания и памяти. Так, группа ОНР с
280
недостаточностью преимущественно мнестических процессов отличается наибольшей сравнительно и с нормой, и с ОНР/ДВ величиной компонента РЗОО в каудальных (особенно в затылочных и задневисочных) отделах коры левого полушария, что сочетается с наименьшей ДРД (прежде всего в центральной области коры). Слабость реакции активации на новый, впервые предъявляемый сигнал рассматривается как отражение дефицита неспецифических диффузных активирующих влияний со стороны ретикулярной формации ствола мозга, что, как следует из вышеизложенного, в наибольшей степени присуще детям, у которых речевая патология сочетается с выраженными нарушениями процессов запечатления и воспроизведения информации.
Группа с ОНР и дефицитом внимания, напротив, отличается наименьшей амплитудой и ПЛ РЗОО в сочетании с более выраженным (сравнительно с нормой и ОНР/ДП) негативным сдвигом ЭА в период развития РД в большинстве корковых зон, но в первую очередь в лобной и центральной, где указанный характер изменений противоположен выявленной в норме и при ОНР/ДП позитивации ЭА. Можно думать, что анализируемые электрографические показатели ОР отражают разные аспекты функционирования сложной системы, обеспечивающей реализацию данного процесса. Большему пониманию функционального смысла выявленных межгрупповых различий должно способствовать исследование изучаемых показателей в другой экспериментальной ситуации - в условиях привлечения внимания к стимулам.
Б. Условия произвольного внимания. Привлечение внимания к стимулам не вызывает, как и в норме, существенных изменений амплитуды РЗОО у детей с ОНР: отмечается лишь тенденция к его росту в каудальных отделах, особенно в правом полушарии у детей с ОНР/ДП; при этом в лобных областях (а также в левой передневисочной зоне в группе ОНР/ДВ) выраженность РЗОО, напротив, несколько уменьшается, т.е. усиливается свойственная норме негативация ЭА переднеассоциативных зон коры (рис. 7.7, А). Указанные изменения способствуют формированию более четкого фокуса РЗОО в теменных отделах обоих полушарий в группе детей с ОНР/ДВ (р < 0,01 сравнительно с С, F, Та, Тр), а при ОНР/ДП - только в правом полушарии (р < 0,01), тогда как в левом сохраняется преобладание амплитуды РЗОО в затылочной зоне.
281



Рис. 7.7, А. Влияние уровня внимания на характеристики электрокорковых компонентов ОР в различных областях коры левого (L) и правого (R) полушарий мозга у двух групп детей с ОНР (с дефицитом внимания -ОНР/ДВ и дефицитом памяти - ОНР/ДП) в сопоставлении с нормой. А - амплитуда компонента РЗОО (мкВ/мс); Б - пиковая латентность РЗОО (в мс); В - величина сдвига ЭА в период РД (мкВ/мс)
Изменения ПЛ РЗОО при повышении уровня внимания более существенны: она, как и в норме, уменьшается при повышении значимости стимулов (рис. 7.7, Б). Однако в группе ОНР/ДВ изменения выражены в наименьшей степени и проявляются лишь в виде тенденции (особенно в передневисочной зоне левого полушария, а также в теменном и центральном отделах правой гемисферы). При ОНР/ДП (рис. 7.7, В), напротив, изменения ПЛ наиболее генерализованны (ГЭФ ФС: р < 0,007) и отмечаются билатерально как в каудальных (О, Тр), так и в передних (F) отделах (р < 0,05), а также в левой передневисочной и в правой теменной зонах (р < 0,05).
282

Рис. 7.7, Б (продолжение)
283




Рис. 7.7, В (окончание)
В результате указанных сдвигов межгрупповые различия амплитуды РЗОО изменяются и количественно (ГЭФ Объект: р < 0,03), и качественно: они наиболее проявляются не в задне-височной коре (что отмечалось при ненаправленном внимании), а в проекционной (затылочной) и переднеассоциативных (лобная и передневисочная) зонах коры (р < 0,05). При этом усиливается выраженность отличий группы ОНР/ДП от нормы и группы ОНР/ДВ. Так, подчеркивается преобладание амплитуды РЗОО сравнительно с нормой в затылочном и лобном отделах, прежде всего правого полушария (р < 0,05). При срав-
284
нении двух речевых групп также выявляется преобладание РЗОО у детей с ОНР/ДП (ГЭФ Объект: р < 0,01), наиболее выраженное в затылочных и лобных отделах (р < 0,05), а также в передневисочных (чаще слева) и задневисочных (больше справа) областях (р < 0,05; рис. 7.8, А).
Межгрупповые различия ПЛ РЗОО, напротив, сглаживаются. Только группа ОНР/ДП наиболее существенно отличается от других испытуемых большей ПЛ РЗОО, при этом различия особенно выражены в каудальных отделах (сравнительно с нормой - в затылочной коре; сравнительно с ОНР/ДВ - в затылочной и теменной; р < 0,05; рис.7.8, Б). Различия между нормой и ОНР/ДВ, напротив, уменьшаются.
Межзональные различия ПЛ РЗОО у всех детей четко не проявляются (однако при ОНР они выражены еще меньше, чем в норме), при этом наиболее коротколатентный РЗОО регистрируется в каудальных отделах правого полушария (в затылочном — в норме и при ОНР/ДП; в теменном — при ОНР/ДВ), хотя межполушарные различия ПЛ несущественны.
Сопоставление ДРД исследованных областей коры на ир-релевантные и релевантные стимулы также позволяет оценить зависимость уровня корковой активации от значимости поступающей информации. Обнаружено, что в целом по группе детей с ОНР, как и в норме, привлечение внимания к стимулам (повышающее их сигнальную значимость) приводит к уменьшению ДРД в затылочной коре; ДРД центральной области при этом увеличивается (см. табл. 7.2). Однако у детей с ОНР указанное увеличение ДРД в центральной корковой зоне при повышении значимости стимулов выражено в меньшей степени, чем в норме (на 115 и 200 мс соответственно, что может быть обусловлено недостаточностью кортикофугальных регулирующих влияний вследствие незрелости лобных отделов коры головного мозга. Это коррелирует с поведенческими характеристиками (отвлекаемость, неустойчивость внимания), выявляемыми у детей с ОНР при психологическом обследовании.
Наибольшие различия по сравнению с нормой выявляются при дифференциации детей на группы с ДВ и ДП и обнаруживаются у детей с ОНР и нарушениями памяти: изменения ДРД в связи с повышением значимости стимулов практически отсутствуют и в затылочной, и в центральной корковых зонах.
285



Рис. 7.8. Особенности электрокорковых компонентов ОР в различных областях коры левого (L) и правого (R) полушарий мозга у двух групп детей с ОНР (с дефицитом внимания - ОНР/ДВ и дефицитом памяти -ОНР/ДП) в сопоставлении с нормой в условиях произвольного внимания. Среднегрупгювые значения: А - амплитуды компонента РЗОО (мкВ/мс); Б - пиковой латентности РЗОО (в мс); В - величины сдвига ЭА в период РД (мкВ/мс)
286
Следовательно, повышение значимости информации в условиях произвольного внимания (речевая инструкция), способствующее усилению регулирующих воздействий со стороны лобных отделов коры, приводит к закономерному, но менее выраженному, чем в норме, увеличению длительности реакции активации ассоциативных отделов коры только у детей с ОНР и дефицитом внимания.
Таким образом, для детей с ОНР в целом и особенно для группы с мнестическими нарушениями характерна значительно меньшая выраженность реакции активации, связанная, очевидно, с недостаточностью восходящих активирующих воздействий и кортикофугальных регулирующих влияний на мезоди-энцефальные структуры головного мозга вследствие незрелости или неполноценности переднеассоциативных отделов коры, фронто-таламических связей.
Обнаружены также межгрупповые различия региональной специфики выраженности электрографических проявлений ОР и их изменений при повышении уровня внимания. При ОНР, сочетающимся с дефицитом внимания, как и в норме, повышение значимости стимулов приводит к изменению на противоположное соотношение степени вовлечения проекционных и ассоциативных областей коры: если в условиях непроизвольного внимания ДРД в затылочной коре превышает таковую в центральной области (особенно у детей с ОНР в связи с существенно меньшей ДРД в центральной коре), то при направленном внимании значения ДРД в центральной зоне коры значительно превышают таковые в затылочной.
Напротив, у детей с ОНР, сочетающимся с недостаточностью мнестических процессов, отсутствует характерное для нормы и для детей с нарушениями внимания преимущественное вовлечение ассоциативной коры (по сравнению с проекционной) в анализ значимой информации. Именно у детей этой подгруппы ДРД центральной коры на релевантные стимулы значительно меньше (на 240 мс) таковой в затылочной коре (см. табл. 7.2).
Изменения характеристик ЭА в период развития РД при повышении уровня внимания также менее существенны, чем в норме. При этом в группе ОНР/ДВ несколько уменьшается негативация переднеассоциативных, фронтальных отделов коры; негативация левой передневисочной зоны, напротив, несколько возрастает, что формирует асимметрию этого показателя (р < 0,05). При ОНР/ДП изменения наименее выра-
287
жены: отмечается лишь тенденция к усилению негативации ЭА в правой затылочной области (рис. 7.7, В). В результате этих сдвигов различия между нормой и ОНР/ДВ по данному показателю становятся несущественными. Напротив, отличия ОНР/ДП от нормы усиливаются в сравнении с ситуацией непроизвольного внимания (ГЭФ Объект: р < 0,05), что проявляется в существенно большей негативации ЭА в ассоциативных (передних /F,C/ и задних /Р/) отделах в норме (р < 0,05; рис. 7.8, В).
Различия между группами с речевой патологией также несколько видоизменяются: при сохранении большей негативации ЭА у детей с ОНР/ДВ (ГЭФ Объект: р < 0,05) различия уменьшаются в правом полушарии (особенно в лобной зоне), тогда как в левой передневисочной области коры они, напротив, возрастают (р < 0,05; рис. 7.8, В).
Изложенное выше подтверждает, что в обеих экспериментальных ситуациях наибольшие межгрупповые различия РЗОО выявляются в проекционных (затылочных) отделах и проявляются в преобладании его амплитуды (и ПЛ в ситуации направленного внимания) у детей с нарушениями речи и памяти, тогда как у детей с дефицитом внимания эти показатели наименьшие. Различия РД, напротив, наиболее выражены в пе-реднеассоциативных зонах, причем при восприятии новых, неожиданных сигналов в условиях непроизвольного внимания в наибольшей степени отличаются от нормы дети группы ОНР/ДВ, тогда как при восприятии значимых стимулов в условиях направленного внимания наибольшие различия выявляются между нормой и группой ОНР/ДП.
У детей с ОНР и дефицитом внимания специфика сравнительно с нормой проявлений ОР заключается в меньшей амплитуде и ПЛ компонента РЗОО, менее продолжительной РД и в проекционных, и в ассоциативных зонах, что сочетается с избыточной, генерализованной негативацией корковых структур, прежде всего переднеассоциативных отделов коры. Повышение уровня внимания способствует, главным образом, увеличению длительности РД и уменьшению негативации ЭА в период ее развития (особенно в лобных отделах), при этом несколько усиливается и компонент РЗОО в каудальных отделах. Это приводит к сглаживанию различий между нормой и детьми с дефицитом внимания.
Следует подчеркнуть, что отличия от нормы исследованных характеристик ОР у детей с ОНР в структурах правого по-
288
лушария выявляются преимущественно в ситуации непроизвольного внимания, т.е. при восприятии новых стимулов. При этом группа ОНР/ДВ отличается от нормы главным образом по характеристикам сдвига ЭА в период РД, а группа ОНР/ДП - по амплитуде РЗОО. Специфика вовлечения левого полушария (его ассоциативных отделов) проявляется при повышении уровня внимания только у детей с ОНР/ДП. Эти данные, очевидно, указывают на особенности участия полушарий в реализации процессов восприятия даже простых невербальных стимулов у детей с ОНР и нарушениями памяти сравнительно с нормой.
Сравнение степени вовлечения проекционных и ассоциативных отделов коры также позволило выявить ряд особенностей, указывающих на специфику нейрофизиологических механизмов ОР при нарушениях речевого развития. Амплитудный максимум компонента РЗОО у детей всех групп отмечен в теменных отделах правого полушария (в норме это проявлялось и в левом) в обеих экспериментальных ситуациях, при этом межзональные различия его амплитуды наиболее выражены в норме, а наименее - в группе с нарушениями памяти. В соответствии с имеющимися в литературе многочисленными данными о связи РЗОО с различными внесенсорными факторами (новизна, значимость стимула, уровень внимания), можно думать, что у детей с ОНР/ДП их влияние реализуется менее специализированно, через более "распределенную" систему связей, при относительной недостаточности вовлечения заднеассоциативных отделов (особенно левого полушария), что может быть одной из причин неполноценности процессов восприятия информации.
Несколько иные межзональные соотношения выявлены для характеристик реакции активации. У детей с нарушениями внимания, как и в норме, сопоставление значений ДРД в затылочной и центральной областях коры позволяет обнаружить, что центральные, ассоциативные зоны вовлекаются более интенсивно, чем затылочные, в процесс анализа значимой информации. Напротив, у детей с дефектами памяти центральные области во всех ситуациях активируются слабее затылочных и не отмечается существенных динамических изменений реактивности центральных отделов коры при повышении значимости стимулов. Это коррелирует и с отсутствием негативации переднеассоциативных отделов в период развития РД в обеих ситуациях.
289
Полученные результаты показывают, что, несмотря на значительный полиморфизм состава детей с ОНР, обусловленный различными соотношениями и сочетаниями степени выраженности и характера нарушений речевого развития и развития познавательной деятельности, возможно выделение ряда существенных отличительных особенностей интегративной деятельности мозга при речевой патологии. Психо- и нейрофизиологические характеристики уровня зрелости корковых механизмов регуляции, степени реактивности различных областей коры и состояния лимбико-диэнцефальных, мезенцефальных и корковых структур, проявляющиеся в покое и при ОР, с учетом клинических и психолого-педагогических данных могут способствовать пониманию структуры дефекта при ОНР и иметь дифференциально-диагностическое значение. Результаты исследования свидетельствуют о том, что для повышения эффективности обучения необходимо оптимальное, адекватное структуре дефекта сочетание логопедической работы и специальных, целенаправленных педагогических приемов, учитывающих специфику нарушений развития когнитивных процессов. Учебная деятельность детей с ОНР, у которых отмечается выраженная нормализация процессов активации мозговых структур при повышении эмоциональной и познавательной значимости предъявляемой информации, может быть значительно улучшена с помощью педагогических приемов, способствующих оптимизации процессов корковой регуляции. Коррекционная работа с детьми, у которых не наблюдается существенной реорганизации степени вовлечения в ОР различных корковых зон, в особенности ассоциативных отделов коры при восприятии релевантной информации, должна быть направлена прежде всего на повышение эффективности ассоциативных процессов, лежащих в основе формирования условных связей, запечатления и воспроизведения информации.
7.5. Специфика процессов регуляции
и следообразования у детей с ЗПР и ОНР
в аспекте проблем дифференциальной
диагностики когнитивных нарушений
Проведенное нами исследование фунционального состояния ЦНС, особенностей восприятия сенсорной информации различной значимости у детей с ЗПР и ОНР сравнительно с нормой позволило выявить совокупность электрофизиологических показателей, анализ которых способству-
290
ет более глубокому пониманию нейрофизиологических механизмов нарушений познавательной деятельности при данных аномалиях развития.
Установлено, что важной отличительной особенностью активности мозга значительной части детей с ЗПР и ОНР в состоянии покоя является недостаточная сформированность ритмики альфа-диапазона, что проявляется в ее нерегулярности, дезорганизации, замедленности, поличастотности. Такие особенности, в соответствии с литературными данными о более позднем формировании в онтогенезе высокочастотных составляющих ЭА коры мозга (Структурно-функциональная организация..., 1990), могут быть связаны с незрелостью генераторов альфа-ритма. Это, в свою очередь, может обусловливать снижение эффективности познавательной деятельности, поскольку несформированность ритмической активности альфа-диапазона рассматривается как отражение слабого развития циклических процессов в ЦНС, через которые реализуются запись поступающей информации и ее считывание, и как коррелят низкого уровня ее пластичности (Дж. Эделмен, 1981; М. Верцеано, 1982; Г.А. Вартанян и др., 1981; Л.И. Переслени, 1984; С.С. Бекшаев и др., 1988). В ряде исследований также выявлена положительная корреляция частотно-амплитудных характеристик альфа-активности с успешностью обучения, с эффективностью когнитивных аспектов деятельности, с уровнем интеллекта (в частности, вербального) (М.Н. Фишман, 1989; И.П. Лукашевич и др., 1994; Th. Gasser et al., 1983; D. Giannitrapani, 1985). В этом контексте существенным представляется тот факт, что указанные особенности ритмики альфа-диапазона наиболее часто отмечались у детей с нарушениями мнестических процессов, причем не только в младшем, но и в старшем школьном возрасте. Эти данные, свидетельствующие о слабой возрастной динамике уровня развития альфа-активности, указывают, на наш взгляд, скорее на неполноценность механизмов его генерации, чем на их незрелость.
У детей с трудностями запечатления и воспроизведения информации наиболее часто выявлялись и знаки локального повреждения корковых структур, преимущественно теменно-за-тылочно-височных отделов правого полушария.
Для детей с ЗПР и ОНР характерна также значительная частота встречаемости и большая, чем в норме, степень выраженности ЭЭГ-знаков, указывающих на отклонения в функциональном состоянии стволовых регуляторных структур
291
мозга. При этом в подгруппах с ДП превалировали знаки дисфункции диэнцефальных (в особенности верхнестволовых) структур мозга, тогда как для детей с ДВ более характерной была дисфункция нижних отделов ствола мозга. Эти данные могут служить дополнительным свидетельством в пользу предположения о неполноценности у выделенных нами подгрупп детей с ДП преимущественно нейрофизиологических механизмов мнестических процессов: разнообразные нарушения памяти при повреждении лимбико-диэнцефальных и ме-зэнцефальных структур головного мозга широко описаны в литературе (А.Р. Лурия, 1974; Н.П. Бехтерева, 1985; Н.К. Корсакова, Л.И. Московичютс, 1985).
Предпринятое нами изучение структурно-функционального обеспечения ОР при восприятии зрительной невербальной информации у детей с ЗПР и ОНР предоставило информацию об особенностях интегративной деятельности мозга при данных аномалиях развития. Исследование активности структур мозга при восприятии информации в условиях непроизвольного внимания и ее реорганизации при повышении уровня внимания позволило проанализировать не только степень сформированное™ неспецифических активирующих влияний, но и особенности корковой регуляции. Это весьма важно в аспекте проблем диагностики когнитивных нарушений, поскольку дает возможность оценить состояние, степень сформированно-сти нейрофизиологических механизмов, лежащих в основе организации и регуляции познавательной деятельности. Сопоставление выраженности различных ЭЭГ-коррелятов ОР в проекционных и ассоциативных зонах коры, вовлекаемых в обработку сенсорной информации в условиях "пассивного" восприятия и в ситуации активного анализа сигналов (при подсчете их количества), позволяет оценить специфику участия этих отделов коры в процессах обнаружения и идентификации новой, неожиданно поступающей информации в ситуации непроизвольного внимания и при отслеживании значимых сигналов в условиях направленного внимания.
Сравнительный анализ характеристик ОР у детей с речевой патологией и с ЗПР выявил значительное их сходство, которое в основном проявляется в слабом по сравнению с нормой вовлечении в реализацию ОР и проекционных, и ассоциативных областей коры головного мозга. Как известно, реакция десинхронизации корковой ритмики на новый, впервые предъявляемый сигнал возникает вследствие неспеци-
292
фических диффузных активирующих влияний со стороны ретикулярной формации ствола мозга на кору. Меньшую, чем в норме, продолжительность данной реакции у всех детей с ЗПР и ОНР, очевидно, можно рассматривать как указание на недостаточность активационных процессов, связанных с реализацией РД. Это предположение подкрепляется выявленными у значительной части детей с ЗПР и ОНР при анализе ЭЭГ признаками заинтересованности нижнестволовых структур мозга, включающих ретикулярные образования продолговатого мозга и моста.
Наиболее важным в аспекте проблем диагностики когнитивных нарушений является тот факт, что характеристики компонентов ОР обнаружили большую корреляцию не со спецификой отклонений в развитии (ЗПР или ОНР), а со структурой нарушений познавательной деятельности - недостаточностью преимущественно процессов внимания или памяти. При этом весьма информативным дифференциально-диагностическим критерием является такой динамический показатель, как изменение соотношения степени вовлечения в ОР проекционных и ассоциативных зон коры при повышении уровня внимания. Установлено, что реакция десинхронизации на новый сигнал в условиях непроизвольного внимания выражена в наибольшей степени в затылочных, специфических отделах коры у всех исследованных групп детей. В ситуации привлечения внимания к таким же по физическим параметрам световым стимулам в норме происходит существенное усиление реакции десинхронизации в центральных, ассоциативных областях и уменьшение ее выраженности в проекционных отделах коры головного мозга. Это обусловлено, с одной стороны, угашени-ем диффузной активации по мере формирования "нервной модели" стимула, утраты стимулом качества новизны, а с другой - усилением регулирующих влияний со стороны передне-ассоциативных отделов коры, обеспечивающих локальную активацию, вовлечение только необходимых для решения избранного типа задач систем внутрикорковых и корково-под-корковых связей (Л.П. Латаш, 1968; Е.Н. Соколов, 1969; Н.В. Дубровинская, 1985; Б.И. Котляр, 1989; Структурно-функциональная организация..., 1990; Л.И. Переслени, Л.А. Рожкова, 1990, 1993, 1996; Р. Наатанен, 1998; R. Naatiinen, 1986). Такие изменения рассматриваются как отражение формирования преднастройки, соответствующей определенному вероятностному прогнозу, которая обеспечивает более точ-
293
ную, экономную реакцию на стимул (И.М. Фейгенберг, 1998). Характерное для нормы увеличение ДРД в центральной зоне коры в условиях произвольного внимания отмечалось только у детей с ДВ. При этом у детей с ЗПР младшего школьного возраста ДРД возрастала также и в проекционной, затылочной коре, указывая на более генерализованное, чем в норме, усиление активационных процессов в связи с повышением значимости воспринимаемой информации, что в некоторой степени компенсирует их недостаточность, наблюдаемую в ситуации непроизвольного внимания.
Сопоставление значений ДРД в затылочной и центральной областях коры позволило обнаружить, что только у детей в норме и с нарушениями внимания центральные, ассоциативные зоны коры вовлекаются более интенсивно, чем затылочные, в процесс анализа значимой информации. Следует отметить, что в норме такое соотношение ДРД, свидетельствующее о преимущественной активации ассоциативных зон коры по сравнению с проекционными, отмечалось во всех возрастах, но особенно выражено было у подростков. Это, очевидно, является отражением присущего данной ситуации дефинитивного паттерна активации мозговых структур.
Изучение ряда других показателей ОР (амплитудно-временные параметры компонента РЗОО, величина и полярность сдвигов ЭА в период развития РД) у детей с ОНР в сопоставлении с нормой позволили получить ряд дополнительных сведений, подкрепляющих высказанные предположения. Так, в подгруппе детей с ОНР и нарушениями преимущественно функции внимания выявлена меньшая, чем в норме и у детей с ДП, амплитуда компонента РЗОО, что также может отражать недостаточную интенсивность активационных нейрофизиологических процессов, определяющих величину РЗОО. Сходные результаты, указывающие на меньшую амплитуду позитивных компонентов с ПЛ в интервале 300 мс у детей с дефицитом внимания, получены и в ряде других исследований (Ю.Д. Кропотов и др., 1999; P. Robaye et al., 1992; B.G. Winsberg et al., 1993). Ю.Д. Кропотов с соавторами (1999) на основании собственных и литературных данных высказывают предположение, что нейробиологической основой синдрома ДВ является дисфункция базальных ганглиев, связанных через тала-мус с корковыми структурами и играющих важную роль в механизмах селекции действий, мыслительных операций. Нарушения этих механизмов поведенчески проявляются в невни-
294
мательности, трудностях удержания внимания на конкретном деле, патологической отвлекаемости, в трудностях подавления поведения, что часто сопровождается гиперактивностью. Сходных взглядов на механизмы дефицита внимания придерживаются и другие исследователи (К.К. Voeller, 1991; E.L. Harris et al., 1995). В этом аспекте интересными представляются результаты, полученные V.R. Brever с соавторами (1997), которые подчеркивают, что дети с гиперактивностью, имея интактную способность переключать внимание, неспособны длительное время удерживать фокусированное внимание, но эта неспособность проявляется только при наличии множества дистракторов, т.е. отвлекающих факторов. Такие особенности функции внимания авторы ассоциируют с недостаточностью передней, фронтальной системы внимания, в отличие от задней системы, дефицитарность которой коррели-руется с нарушениями такого компонента ОР на стимулы, как отключение от незначимых событий с тем, чтобы переключиться на более значимые. Следует отметить, что некоторые из указанных поведенческих особенностей (повышенная от-влекаемость, неустойчивость внимания, двигательная растор-моженность) были присущи и детям, включенным нами в группы с ДВ. Однако в наших исследованиях у этих детей наряду с негрубыми изменениями ЭА верхнестволового генеза отмечались признаки дисфункции каудальных отделов ствола мозга, которые по частоте встречаемости и по степени выраженности превалировали над первыми.
Важным для понимания нейробиологической природы выявленной у детей с ДВ недостаточности активационных процессов является тот факт, что при исследовании детей с ЗПР/ДВ обнаружена положительная возрастная динамика выраженности компонентов ОР. Это позволяет предположить, что в основе наблюдаемых у этих детей нарушений лежит незрелость механизмов активации и корковой регуляции, которая в большей или меньшей степени компенсируется по мере морфофункционального созревания мозговых структур и их связей.
Вместе с тем при нарушениях внимания у детей с ОНР обнаружена большая, чем в норме (а также в группе ОНР/ДП), негативация структур мозга в период развития РД, что может служить электрографическим коррелятом отмечаемой в структуре синдрома ДВ чрезмерной выраженности ряда проявлений ОР на внешние раздражители. Вовлечение в процесс
295
восприятия переднеассоциативных зон коры при повышении уровня внимания уменьшает степень негативации, что, очевидно, отражает некоторую нормализацию активационных процессов. Уменьшение "избыточной" негативации зрительных ВП, присущей части детей с дефицитом внимания и гиперактивностью, отмечается под влиянием метилфенидата, что рассматривается как отражение нормализации процессов активации (Y. Frank, 1993). Любопытными в этом аспекте представляются также данные, свидетельствующие о негативном сдвиге уровня постоянного потенциала головного мозга у детей с гиперактивным поведением (в отличие от характерного для нормы позитивного сдвига), что рассматривается как коррелят снижения общего уровня энергозатрат, обеспечивающих нейрофизиологические механизмы регуляции жизнедеятельности (Н.Г. Городенский, С.Л. Шармина, 2000). По-видимому, эти процессы также могут находить отражение в выявленных нами особенностях ОР, влияя и на характер сдвига"ЭА в период развития реакции активации, и на амплитуду позитивного колебания РЗОО (уменьшение которой наблюдается у детей с ДВ).
Иная специфика реализации ОР выявляется у детей с нарушениями преимущественно мнестических процессов. Обнаружено, что в группах с ДП центральные области во всех ситуациях (а при ЗПР и во все возрастные периоды) активируются слабее затылочных и не отмечается существенных динамических изменений реактивности центральных отделов коры при повышении значимости стимулов. У детей с ОНР это коррелирует и с отсутствием негативации переднеассоциативных отделов в период развития РД в обеих ситуациях.
В соответствии с литературными данными в центральных, ассоциативных отделах коры в отличие от зрительных и слуховых областей широко представлены полимодальные нейроны, что обеспечивает формирование ассоциативных (или условных) связей между сенсорными стимулами и адекватными поведенческими реакциями. Способность формировать ассоциативные связи рассматривается как главный признак памяти; при их образовании происходит ряд последовательных событий, завершающихся структурной модификацией межнейронных связей, что в итоге проявляется в изменении активности клеток (Е.А. Громова, 1980; С.- Роуз, 1995; К.В. Анохин, 1997). Выявляемая у всех детей,с ЗПР и ОНР недостаточность вовлечения центральных областей коры может рассматри-
296
ваться как одна из причин слабости ассоциативных процессов. Однако важно то, что только у детей с недостаточностью преимущественно процессов внимания (подгруппы ДВ) при повышении значимости поступающей информации, способствующей усилению корковых регулирующих влияний, наблюдается некоторая нормализация интегративных процессов, в значительной мере связанных со скоростью формирования и прочностью ассоциативных связей.
Эти предположения подкрепляются также данными, свидетельствующими о характерном для нормы закономерном и значительном усилении с возрастом степени активации ассоциативных зон коры при предъявлении релевантной информации, что, на наш взгляд, можно оценивать как отражение расширения связей этих областей с различными структурами головного мозга. Это может способствовать повышению успешности решения сложных перцептивно-мыслительных задач, которое реализуется на основе хранящейся в памяти информации и обеспечивается содружественной работой различных образований головного мозга при актуализации широких, разнообразных внутрикорковых и корково-подкорко-вых связей. При нарушениях памяти у детей с ЗПР, напротив, на протяжении всего исследованного возрастного диапазона (от 7 до 15 лет) отмечается слабое вовлечение в ОР ассоциативной коры. Возможно, это обусловлено неполноценностью, ограниченностью ассоциативных связей, которые с возрастом хотя и обогащаются, но крайне недостаточно, что может быть связано с нарушениями процессов следообразо-вания. Косвенным подтверждением данного предположения является и обнаруживаемое у детей с ЗПР/ДП отсутствие характерного для нормы выраженного декремента ДРД при угашении ОР, которое базируется на ассоциативном механизме мозга (Б.И. Котляр, 1989) и отражает процесс "негативного" научения, связанный с формированием "нервной модели стимула" (Е.Н. Соколов, 1969). В этом аспекте важными представляются современные исследования, в которых механизм консолидации долговременной памяти отождествляется с активацией ряда молекулярных процессов в нервных клетках при обучении (К.В. Анохин, 1997). Показано, что этот механизм является универсальным и проявляется как реакция генома клетки на изменения экстраклеточной среды, которая активируется новизной ситуации и возникновением ориентировочно-исследовательской деятельности и
297
включает последующие процессы морфогенеза (рост и диф-ференцировка клеток), что позволяет рассматривать обучение и развитие мозга как взаимосвязанные процессы. Можно предположить, что выявленная у детей с ДП специфика пространственно-временной организации активационных процессов при ОР (особенно недостаточность вовлечения ассоциативных зон коры при восприятии значимой информации) затрудняет "запуск" этого универсального механизма, в связи с чем нарушаются и процессы морфогенеза, и процессы запечатления информации, а следовательно - и обучение.
Анализ ряда других проявлений ОР при нарушениях памяти у детей с ОНР показал, что для них характерна большая сравнительно с нормой и группой с ДВ выраженность компонента РЗОО в затылочных отделах. Можно предположить, что более интенсивное, чем в норме, вовлечение в ОР проекционной, затылочной зоны коры (высокоамплитудный РЗОО) сочетается с быстрым затуханием активирующих влияний (кратковременная РД), и это обусловливает недостаточное вовлечение в реализацию ОР переднеассоциативных зон коры. Выявленное отсутствие выраженного негативного сдвига ЭА в период развития реакции активации с учетом вышеприведенных данных можно рассматривать как отражение недостаточной активации коры мозга (особенно его лобных отделов при восприятии релевантной информации), обусловленной дисфункцией лимбико-гипоталамического комплекса, наиболее тесно связанного с медленной управляющей системой, а также заинтересованного в обеспечении мнестических функций. Отмечается важная роль гипоталамических активирующих влияний, обеспечивающих более обширное вовлечение проекционной коры, на начальных этапах формирования репрезентативной системы, а также в формировании преднастройки (Е.А. Громова, 1980; А.Я. Могилевский, 1986).
Полученные нами результаты могут быть глубже поняты в аспекте современных представлений о двух иерархически организованных когнитивных системах, системах внимания (A.M. Иваницкий, 1997; Р. Наатанен, 1998; М. Pozner, S. Peterson, 1990). Одна из этих систем, по мнению Р. Наатане-на, представлена сенсорно-специфическими, исполнительными механизмами (направленными на отбор стимулов), а другая - командно-контролирующими механизмами более высокого уровня, что согласуется с классификацией, предложенной М. Познером с соавторами (М. Pozner, S. Peterson, 1990),
298
выделяющими заднюю, париетальную /PAS/ и переднюю, фронтальную /FAS/ системы внимания. С учетом этих представлений результаты, полученные при исследовании группы с ДП, можно рассматривать, по-видимому, как отражение неполноценности преимущественно командной системы, в структуре которой важнейшая роль принадлежит фронто-талами-ческой системе связей. На это указывает и отсутствие достаточно выраженных динамических изменений ЭА (и РЗОО, и РД) при повышении значимости стимулов и именно в перед-неассоциативных зонах коры, и дисфункция преимущественно верхнестволовых регуляторных структур. Передние отделы мозга включаются в так называемую эндогенную систему внимания, которая запускается внутренними стимулами, тогда как экзогенное внимание, связанное с внешними сигналами, вовлекает задние отделы мозга и подкорковые структуры (A. Rosen et al., 1997). Ввиду взаимовлияния, взаимообусловленности аттенционных и мнестических процессов, а также важной роли фронтальных отделов коры в обеспечении мнестических функций (А.Р. Лурия, 1973; М.А. Wheeler et al., 1995) недостаточная интенсивность и длительность реактивности переднеассоциативных отделов коры у детей с ОНР/ДП может обусловливать слабость процессов следообразования, лежащих в основе механизмов запечатления информации.
В связи с изложенным полагаем, что отсутствие существенной реорганизации степени активации различных областей коры при повышении значимости воспринимаемых стимулов (характерное только для детей с нарушениями мнестических процессов) может служить информативным показателем для оценки биологических предпосылок, резервов в развитии некоторых высших психических функций при аномалиях развития и иметь прогностическое значение.
Таким образом, наиболее выраженные нарушения инте-гративной деятельности мозга отмечаются у детей с ДП. При этом, как показали наши исследования, для детей данной группы характерно преимущественное вовлечение в патологический процесс правого полушария, что проявляется в существенно большей частоте встречаемости локальных изменений ЭА в правополушарных структурах (преимущественно затылочно-теменно-височных) сравнительно с ле-вополушарными. Закономерным в связи с этим представляется выявленное у детей с ДП преобладание признаков дисфункции верхнестволовых структур мозга, поскольку из-
299
вестны данные о функциональной связи диэнцефальных структур с правой гемисферой (Н.Н. Брагина, Т.А. Доброхотова, 1981; Г.Н. Болдырева и др., 1997; П.В.Симонов, 1997). Большая уязвимость, чувствительность к различным вредным факторам именно правого полушария на ранних стадиях онтогенеза, по мнению ряда исследователей, может быть обусловлена его морфологическими особенностями — преобладанием в правополушарных структурах сравнительно с левополушарными белого вещества, что предполагает большее количество межрегиональных связей; поэтому если формирование этих связей, коррелирующее с развитием сети дендритов и миелинизацией волокон в раннем онтогенезе, по каким-либо причинам прерывается или задерживается, то это в большей степени сказывается на структурах правого.полушария (М. Semrud-Clikeman, G.W. Hynd, 1990). Среди факторов, обусловливающих структурно-функциональную недостаточность правого полушария, отмечаются отклонения в функциональном созревании мозга, наличие мозговых повреждений (вследствие пренатальных интоксикаций, асфиксии, дегидратации, травмы головы и др.) (Ч. Ньокиктьен, 1994; S.R. Batler, 1997). Важным фактором, определяющим особенности морфофункциональной организации мозга, которые могут обусловливать и специфику протекания высших психических функций, является генетическая детерминированность анатомо-физиологических особенностей мозга и нервной системы. Так, например, отсутствие гена "правостороннего сдвига" рассматривается как возможная причина нарушения полушарного доминирования (Ч. Ньокиктьен, 1994; Т.М. Марютина, 1999); при этом отмечается, что большая генетическая обусловленность характерна для ЭА правого полушария, особенно для его задних отделов (С.Б. Малых, 1997).
Аномальный характер развития структур правого полушария, очевидно, не может не сказываться на формировании познавательных способностей, в том числе лингвистических, поскольку правое полушарие является базовым для развития многих высших психических функций (В.Д. Труш, М.Н. Фиш-ман, 1991; Функциональная асимметрия... 1992; М. Semrud-Clikeman, G.W. Hynd, 1990). Во многих исследованиях с использованием методов позитронно-эмиссионной томографии (ПЭТ) и ядерно-магнитного резонанса (ЯМР) выявлена значительная корреляция речевых нарушений с различными мор-
300
фологическими аномалиями, проявляющимися в отклонениях от нормального паттерна асимметрии мозговых структур (например, отсутствие преобладания височно-теменно-затылоч-ных областей в левом полушарии по сравнению с правым при дислексии) (A.M. Galaburda, 1989; G.W. Hynd, M. Semrud-Clikeman, 1989; G.W. Hynd et al., 1990, 1993, 1995; C.M. Leonard et al., 1993); отмечается также вовлечение в патологический процесс фронтальных отделов преимущественно правого полушария (G.W. Hynd, M. Semrud-Clikeman, 1989). Поражения правого полушария вызывают различные когнитивные нарушения (ухудшение памяти, снижение интеллекта, нарушения внимания) , которые вкупе обозначаются как "трудности научения", в основе которых лежит неспособность использовать сохранные перцептивные и моторные способности, интегрируя их в адекватном новой ситуации способе функционирования (I. Miard, J. Blanc-Garin, 1986).
Таким образом, анализ компонентов ОР при предъявлении индифферентных и релевантных сигналов позволяет выявить особенности по сравнению с нормой интегративной деятельности мозга детей с ЗПР и ОНР в процессе восприятия различной по значимости сенсорной информации, характер которых свидетельствует о некотором сходстве базисных нейрофизиологических механизмов, определяющих ряд нарушений познавательной деятельности у этих категорий детей. Эти особенности с наибольшей отчетливостью проявляются при анализе соотношения степени активации проекционных и ассоциативных отделов в зависимости от значимости поступающей информации и могут способствовать дифференциации детей с ЗПР и ОНР на подгруппы с учетом преимущественной выраженности аттенционных или мнестических нарушений. Совокупность данных, полученных при исследовании этих детей, особенно с превалированием нарушений процессов памяти (меньшая сравнительно с нормой длительность реакции десинхронизации корковой ритмики при предъявлении сенсорных сигналов; слабая выраженность или отсутствие характерного для нормы перераспределения активации кау-дальных и ростральных отделов коры при повышении значимости раздражителей, приводящего к преимущественному вовлечению в ОР непроекционных ассоциативных корковых зон) свидетельствует о существенных отличиях структурно-функционального обеспечения процесса восприятия информации у детей с ЗПР и ОНР.
301
Итак, анализ особенностей ОР при восприятии сенсорных стимулов различной сигнальной значимости в сопоставлении с характеристиками ЭЭГ в состоянии спокойного бодрствования у детей с ЗПР и ОНР дает возможность судить о причинах недостаточности аттенционных и мнести-ческих процессов, выявляемых в ходе психолого-педагогического обследования. Необходимо подчеркнуть, что использование двух ситуаций - ненаправленного и направленного внимания - позволяет обнаружить своеобразие корко-во-подкорковых взаимодействий и те отличия, которые специфичны для испытуемых с разной структурой нарушений познавательной деятельности.
Одна из причин отклонений в развитии высших психических функций - недостаточность активирующих влияний в системах корково-подкорковых связей, а также незрелость процессов корковой регуляции, что лежит в основе дефицита внимания у детей подгруппы ДВ.
Второй причиной снижения эффективности познавательной деятельности и отклонений в характеристиках ОР у детей с ЗПР и ОНР, особенно в случаях его сочетания с дефектами памяти, может быть недостаточная эффективность формирования ассоциативных связей. В результате этого затрудняется образование "нервных моделей" или репрезентаций взаимосвязанных во времени стимулов, без чего невозможно установление зависимостей между событиями окружающей среды, а следовательно - обучение и целенаправленное адаптивное поведение.
Особенности морфофункциональной организации головного мозга при нарушениях памяти у детей с ЗПР и ОНР, проявляющиеся в недостаточном развитии корковой ритмики (меньшая сравнительно с нормой выраженность альфа-активности, специфика ее зонального распределения), а также высокая частота встречаемости локальных изменений ЭА (преимущественно в правополушарных структурах) в сочетании с дисфункцией стволовых регуляторных (особенно верхнестволовых) структур мозга могут быть важной причиной недостаточной сформированности процессов записи и считывания информации.